Prediction 5.1: 유전적 변화
유전정보는 유기체의 모든 것과 그것의 가능성을 자세히 알려줍니다. 유전형(genetype)은 가능한 표현형을 specifiy하며 그렇기 때문에 유전형이 변화하면 표현형의 변화가 뒤따릅니다. 이것은 진화론적인 변화가 보여지는 영영죽의 하나임이 분명합니다. 유전적 변화는 진정으로 진화과정을 이해하기 위해서 가장 중요한 것이다.
확인:
극히 extensive 한 유전적 변화가 실험실이나 야생에서 모두 관찰되었습니다. 우리는 Genetic flow, 무작위적 유전자 drift, 자연선택, 그리고 돌연변이를 포함하는 수 많은 자연 현상에 의해서 게놈이 비가역적으로 바뀌고 유전됩니다. 관찰된 돌연변이는 "movile intrones, 유전자 복제, 재조합, 트랜스토지션, 레트로바이러스 삽입(수평적, 유전자 이동), 염기 결실, 염기 삽입, 그리고 염색체 재배열 (rearragements) 염색체 재배열에는 게놈 복제(eg. 다배체성) 불균형 교차, 역위, 위치전환(전좌), 분열(fissions), 유전자 fusions, 염색체 복제 및 염색체 결실이 포함됩니다 (Futuyma 1998, pp. 267-271, 283-294). .
반증가능성
유전자 물질이 발혀지자, 대진화가 진행되기 위해서는 유전물질의 많은 변화가 필요한 것이 분명합니다. 만약 유전학자들이 유전자를 관찰하여 게놈 정지상태나 혹은 변화에 저항한다면 대진화에 대한 중요한 반대증거가 될 것입니다. 중요한 유전자 변화에 대해서 예를들어, 우리가 어떤 유기체를 돌연변이 시켰을 때, 그 DNA가 다시 되돌연변이하여 전 상태로 돌아올 수도 있을 것입니다. 그러나, 실제로는 그 반대입니다. 게놈은 믿을 수 없이 유연하며 유전적인 변화는 유전되고 본질적으로 비가역적입니다(Lewin 1999).
예측 5.2: 형태학적 변화
분지학적 분류, 그리고 계통발생적인 재구성은 주로 다양하고 쉽게 구분되는 종들의 형태학적 특징에 근거하고 있습니다. 대진화는 진화사를 통한 유기체의 형태의 변화가 필요합니다. 그러므로 우리는 현재의 집단에서도 형태학적인 변화와 변이를 관찰할 수 있을 것입니다.
확인 :
유기체 집단에서 형태학적 변화의 수많은 관찰을 해왔습니다(Endler 1986). 이러한 변화에는 일부 장기의 색의 변화 예를들어 초파리의 노란색 몸과 갈색군, 생쥐의 털색(Barsh 1996), 물고기의 비늘색(Houde 1988), 새의 깃털 패턴(Morton 1990)과 같은 사례가 있습니다. 동물들의 크기, 길이, 너비, 혹은 신체적인 특징등의 모든 상상할 수 있는 유전적인 변이가 기록되어 있습니다(Johnston and Selander 1973; Futuyma 1998, p. 247-262). 이들 중 마지막 사실은 공통조상에는 매우 중요한 것입니다. 왜냐하면 많은 종들에게 있어서 주요한 형태학적 변화들은 (예를들어 양서류 파충류, 포유류와 조류들의 종들은) 각각 해당하는 상동기관 구조의 어떤 부분의 크기의 변화일 뿐이기 때문입니다.
예측 5.3: 기능적인 변화
유기체 들의 주요한 차이의 하나는 다양한 기능의 그들의 capacity입니다. 한 생태학적 지위(niche)를 차지하기 위한 능력은 변함없이 기능을 다르게 하는 것입니다. 그러므로 기능의 변화는 대진화적 변화에 매우 중요합니다.
확인:
많은 유기체는 전에는 가지고 있지 않은 새로운 기능을 획득하는 것이 관찰되었습니다(Endler 1986). 박테리아는 바이러스 (Luria and Delbruck 1943)와 항생제에 대한 저항성(Lederberg and Lederberg 1952)을 획득했습니다. 박테리아는 또한 새로운 아미노산과 핵산을 합성하는 능력을 진화시켰습니다(Futuyma 1998, p. 274). 단세포 유기체는 자연에는 없었던 인공 화학물질인 나이론이나 펜타클로로페놀만을 유일한 탄소원으로 사용할 수 있는 능력을 진화시켰습니다 (Okada et al. 1983; Orser and Lange 1994). 특히 이중에 뒤의 능력을 획득하는 것은 전혀 새로운 멀티효소 대사과정을 진화시켜야 합니다(Lee et al. 1998). 박테리아는 이전에는 살수 없는 온도에서 성장하도록 진화되어 왔습니다(Bennett et al. 1992). 대장균에서는 우리는 완전히 새로운 대사게의 진화를 볼 수 있습니다. 새로운 탄소원을 대사하는 능력, 새로운 조절유전자에 의해서 이 능력을 조절하는 능력, 세포막을 통해서 새로운 탄소원을 전달시키는 능력들이 포함됩니다 (Hall 1982).
이러한 새로운 기능의 진화론적인 획득은 후생동물에서도 흔합니다. 우리는 곤충들이 살충제에 저항성이 생기는 것(Ffrench-Constant et al. 2000), 동물과 식물이 병에 대한 저항성을 갖는 것(Carpenter and O'Brien 1995; Richter and Ronald 2000), 갑각류의 포식자에 대한 방어를 새로 진화시키는 것(Hairston 1990), 양서류는 서식처의 산성화에 저항력을 갖도록 진화하는 것(Andren et al. 1989), 포유류가 독에 대한 저항성을 획득하는 것(Bishop 1981)을 관찰할 수 있습니다.
공통조상 이론으로부터 얻어지는 가장 일반적인 결론은 생명체 전체가 과거에는 달랐다는 것입니다. 우리가 좀 더 오래전 과거를 돌아보면 볼수록 현재의 생명계와는 좀 더 다른 생명체들을 볼 수 있습니다. 좀더 최근의 화석들은 오래된 화석보다 훨씬 현재와 비슷합니다.
이점은 공통조상을 예측한 1.4와 1.5와는 약간 다른 것입니다. 우리가 보아왔듯이 계통발생수는 많은 공통조상과 그들의 형태를 예측해 줍니다. 그러나, 종 역학(species dynamics)과 최근 멸종 비율이 주어진다면 우리는 주요 유기체들이 결국 멸종될 것을 알 수 있습니다 (Diamond 1984a; Diamond 1984b; Wilson 1992, ch. 12; Futuyma 1998, pp. 722-723). 이것을 외삽하면 과거의 유기체 대부분은 이미 멸종되었을 것입니다. 그러므로 우리는 합리적으로, 많은 다른 후손과 친척을 가진 것으로 오늘날까지 살아남은 후손을 가지지 못한 예상 공통조상을 기대할 수 있다. 간단히 말해서 우리는 우리가 발견할 화석 종의 대부분이 현대 종의 실재 공통조상이 아닐 것이며 아마도 결국은 멸종된 친척관계에 있었던 생명체 일 것입니다.
확인:
우리가 발견한 가장 오래된 돌은 41억년 전 것이며 그들은 어떠한 생명체도 가지고 있지 않았습니다. 그후로 20억년 후의 시원대(Archean, 始原代)에서 발견된 돌은 전혀 다세포 생물은 없고 원핵생물만 있었습니다. 21억년 전의 돌에서 최초의 진핵생물(핵을 가진 단세포 생물)이 나타납니다. 다시 10억년이 지나도 다세포 생물에 대한 증거는 없었습니다.처음으로 다세포 유기체의 존재에 대한 힌트가 나타나는 것은 11억년전 사암에서 발견된 작은 벌래(worm, 대개 환형동물류의 벌레를 말함)이 땅을 파고 들어간 (burrow) 흔적 화석이었습니다.
선캠브리안/캠브리안 전환시기 근처인 5억 8천만년 전의 '에디아카라'와 버제스 쉐일 (Burgess shale)동물상에서 우리는 다세포 생물의 첫 번째 화석을 발견할 수 있었습니다. 그러나 그들은 매우 이상하고, 작고, 연한 몸을 가진 생물이었으며, 겉으로 보기엔 대부분이 오늘날 발견되는 그 어떠한 것과도 닮지 않았습니다. 표준 계통발생수로부터 기대한 대로 최초의 다세포생물 화석은, 매우 단순한 해면과 말미잘(말미잘과 해파리는 모두 유자포동물(cnidarian)이다.)을 닮은 유기체입니다. 이로부터 2천만년 뒤에 우리는 단순한 연체동물, 벌래 및 극피동물(불가사리나 해삼과 비슷한 유기체)를 발견하게 됩니다. 다시 약 1천5백만년 쯤 지나야 비록 대부분이 사람들이 척추동물이라고 인식하기 힘들겠지만, 가장 최초의 척추동물이 나타나게 됩니다. 그들은 그들은 작은 벌래를 닮았고 원시 물고기를 닮은 유기체로 뼈, 턱, 혹은 지느러미(등지느러미는 제외)가 없었습니다.
원생대를 거쳐 현생으로 오면서 생명체는 현대의 생물상과 좀더 급격하게 비슷해 집니다. 캠브리안 시기에는 (5억 4천 - 5억년) 우리는 예를들어 삼엽충, 해면, 극피동물(echinoderm)과 같은 무척추동물을 주로 발견하게 됩니다. 이 시대에 발견되는 유일한 척추동물인 척색을 가진 벌래를 제외하고는 그 뒤 1억년 동안 바다 생물은 무척추 동물과 이상한 무악어(턱없는 물고기)에 지배됩니다. 우리에게 좀더 친숙한 턱뼈 있는 물고기들은 4억 1만년 전인 실루리아기 후기에 나타납니다. 지구의 지층의 90%인 데본기(4억년)까지 육지동물은 없었습니다.
데본기 동안 우리는 드디어 곤충의 첫 번째 증거를 발견할 수 있습니다. 그리고, 그 뒤로 1억년 동안 석탄기에서 페름기(~3억년까지) 육상에는 파충류, 조류, 포유류가 없었고, 오직 양서류와 곤충만이 있었습니다. 육지는 양치류(fern) 으로 덮혔으며 침엽수나 상수리 나무나 그와 비슷한 어떠한 것도 없었습니다.
중생대 동안(2억 5천 - 6천5백만년) 생명체는 괴물처럼 큰 파충류인 공룡이 지배했습니다. 그 당시 가장 많은 식물은 조금은 이상한 소철과 같은 겉씨식물이었습니다. 약 1억9천만년전 쥬라기가 되기까지 현대 포유류를 닮은 어떠한 것도 발견되지 않았습니다. 1억 9천년에 발견된 이 "포유류"는 작고 반은 파충류 반은 설치류로 아직 거대동물은 나타나지 않았습니다. 지구의 살아있는 유기체의 화석을 포함한 침적물의 90%에는 꽃이 존재한 증거가 없으며, 꽃들은 효신세(Paleocene, 6천5백만년전) 직전에 나타납니다. 이와 마찬가지로 지구의 생명체의 기록은 신생대(6천 5백만전에서 현재까지)이 되기 전에는 단단한 나무 숲이 없었다.
신생대 동안 포유류와 조류가 드디어 오늘날에 보듯이 지구상에 널리 퍼지게 되었으며 충적세(Pleistocene, 2백만년)가 될 때 까지는 지구의 생물상은 완벽하지는 못해도 매우 비슷해지면, 차이가 있다면, 거대동물 매머드, 자이언트 늘보(giant sloth), 검치호와 같은 유기체가 대륙을 덮고 있었다(Futuyma 1998, pp. 130, 169-199).
반증가능성:
이 반증가능성은 아주 간단하고 아주 쉬운 것이다. 지구의 침저거물이 더 오래되면 될수록 현대의 생명체들과 비슷한 종들이 분포하면 된다.
성이 있는 후생동물을 위한 종에 대한 가장 유용한 정의는 생물학적 종 개념입니다(진화를 가정하지 않았습니다.). 즉 종들은 실재적 혹은 잠재적으로 상호교배가능한 자연적인 개체군의 그룹을 말하며 이들은 이와 같은 다른 집단과 생식적으로 분리되어 있어야 합니다(Mayr 1942).
만약 어떠한 종이 새로운 종으로 가지를 치는 과정이 과거에 점차적으로 일어났다면 우리는 오늘날에도 완전히 상호교배가 가능한 것에서부터 일부 상호교배가 가능하고 이후 불임성이 약간 줄어들었거나 혹은 완전히 불임인 상태로 상호교배된 것에서부터 유전적으로 완전히 분리된 개체군에 이르기까지, 종분화 혹은 유전적 분리의 모든 단계를 볼 수 있을 것입니다.
확인:
분명히 다른 종이 비정상적인 상황 혹은 제한된 상황에서 잡종을 형성하는 사례는 수도 없이 많습니다. 예를들어 서유럽 까마귀와 아시아 hooded crow는 아주 좁은 "잡종 지역"을 가지고 있습니다. 또 다른 사례로, Catostomus 속의 빨판물고기의 플랫 강가의 종들은 서로 같이 살기도 하지만 상호교배되는 경우가 드믈게 나타납니다(Futuyma 1998, p. 454). .
부분 혹은 불완전 종분화의 가장 놀라운 사례의 하나는 수 많은 "고리 종(ring species)"입니다. 도룡뇽의 일종인 엔산티나(Ensatina)와 같은 고리종은 어떠한 지리적인 특징을 둘러쌓고 상호 교배가 가능한 개체군의 사슬을 형성하며, 이 두 양끝의 집단이 서로 만나게 되면 그들은 서로 마치 다른 종처럼 행동합니다. 엔사티나의 경우 아종은 캘리포니아 중앙 계곡 주변에 고리를 만들고 있으며 이들 아종은 각각 자유롭게 동,서,북면에서 자유롭게 이종교배된다. 하지만 그들이 남쪽에 살고 있는 종과 공존하게 되면 그들은 잡종을 만들 수 없으며 마치 분리된 종처럼 행동합니다(Moritz et al. 1982; Futuyma 1998, pp. 455-456)
다른 고리 종의 사례는 갈매기 Larus.속입니다. L. argentatus 과 L. fuscus는 원래 영국에서는 분리된 종으로 확인되었다. 그러나 Larus 잡종의 연속적인 고리가 북극 주변의 동쪽에서 서쪽까지 뻗쳐있습니다. 오직 영국에서만 이종교배가 안됩니다.
박새 역시 중앙 아시아의 산 주변에 고리 종을 형성합니다. 이들은 중국 북부를 제외하고는 자유롭게 이종교배가 가능합니다(Smith 1993, pp. 227-230).
많은 종들이 잡종을 만들 수 있지만 결과로 나온 후손은 생식력이 떨어진다. 하나의 사례가 영국 뒤쥐(뒤쥐 속)로 이들의 잡종은 염색체가 다르기 때문에 생식적인 불이익을 가지고 있습니다. Drosophila pseudoobscura의 유타와 캘리포니아 스트레인을 교배시킨 실험실 실험에서도 볼수 있다. 다른 사례는 개구리 Bombina bombina와 Bombina variegata 로 이들의 잡종은 적응력이 떨어집니다(즉, 그들은 성공적으로 생식하지 못합니다.).
많은 다른 종들은 생존은 하지만 불임인 잡종을 생산합니다. 이것은 양서류(Rana 속의 일부 개구리 종)와 말(말과 당나귀의 교배는 불임인 노새가 생깁니다.)의 종류에서 관찰됩니다. 다른 사례는 영원(newt)인 Triturus cristatus 와 T. marmoratus 이들의 잡종은 쌍을 이루지 못한 염색체로 인하여 불임이 된다(Smith 1993, pp. 253, 264).
다른 종들은 성공적인 생식이 가능하기도 하지만 배아발생 단계에서 사망하기도 합니다. 이러한 사례는 개구리 Rana. pipiens 와 R. sylvatica입니다(Futuyma 1998, p. 460). 이 현상은 초파리에서도 관찰되기도 합니다. 다른 사례는 목화 종 Gossypium hirsutum 와 G. barbadense 에서 발견됩니다(Smith 1993; Futuyma 1998, ch. 15 and 16).
반증가능성:
만약 모든 종들이 완벽히 유전적으로 서로 서로 분리되어 있으며, 잡종이 생기지 않는다면 과거 수백만번의 수백반번의 점차적인 종분화를 합리적으로 정당화하기 어려울 것입니다.
표준 계통발생수는 수 없이 많은 종분화 사건을 그림으로 보여줍니다. 각각 공통조상은 또한 최소한 1 사례 이상의 종분화 사건을 나타냅니다. 그러므로 비록 매우 드물지만 우리는 실제 종분화 사건을 관찰할 수 있습니다. 완전히 생식분리가 일어나는데 걸리는 시간은 화석기록과 측정된 돌연변이율로부터 볼 때 야생에서 평균 약 300만년이 걸립니다(Futuyma 1998, p. 510). 결론적으로 자연에서 종분화의 관찰은 가능하지만 극히 드믄 현상입니다. 그러나, 실험실 생명체의 진화속도는 화석기록으로 추론하는 것보다 빠르며 일반실험실에서 종분화를 관찰하는 것이 가능해집니다(Gingerich 1983).
확인:
20세기 초기부터 잡종과 다배체(polyploidization)에 의한 종분화가속씨식물과 양치류 (hemp nettle, 앵초, 무 그리고 양배추와 다양한 양치식물)에서 보였습니다. 그리고 식물에서 관찰된 몇몇 종분화 사건은 hybridization 과 polyploidization과 관계가 없었습니다(예를들어 옥수수나 S. malheurensis).
유전학에서 가장 널리 연구된 유기체의 하나는 초파리 종입니다. 많은 초파리 종분화 사건이 1970년대부터 널리 문서화 되었습니다. 초파리의 종분화는 다른 메카니즘 보다 특히 공간 격리, 어떤 지역에서의 서식지 특화, 구애 행동 변화, 파괴적인 자연석택, bottlenecking populations (founder-flush 실험)에 의한 것입니다.
몇몇 종분화 사건은 집파리, gall former flies, apple maggot flies, flour beetles, Nereis acuminata (벌래의 일종), 모기, 및 다른 다양한 곤충의 실험실 군집에서 보여졌습니다. 녹조류와 박테리아는 선택압력으로 인하여 단세포성에서 다세포성으로 형태학적으로 짧은 간균에서 긴 간균으로 변했기 때문에 종분화가 일어났다고 분류되었습니다.
포유류에서도 종분화가 관찰되었습니다. 지난 500년 동안 마데이라(Madira)의 집의 생쥐에 대해서 단지 지질학적 격리, 유전자 전이, 및 염색체 융합의 결과로만으로 6건의 종분화가 있었습니다. 한번의 염색체의 융합이 인간과 침팬지의 유일하고 주된 유전적 차이이며, 마데이라 생쥐들은 지난 500년동안 9번의 염색체 융합에도 살아났습니다 (Britton-Davidian et al. 2000).
좀더 자세하고 많은 참고문헌을 원하는 사람들은 Observed Instances of Speciation FAQ을 참고하기 바랍니다.
예측 5.7: 형태학적인 변화의 속도
진화적인 변화에 대하여 관찰된 속도는 화석기록으로 관찰된 속도보다 빠르거나 같아야 합니다.
확인:
이 문제에 대해서 죠지 윌리암스의 글을 인용하는 것보다 더 나은 것을 없을 것입니다:
1983년 필립 깅그리치는 512건의 서로 다른 진화의 관찰 속도에 대한 분석한 유명한 연구를 출판했습니다(Gingerich 1983). 그 연구는 (1) 실험실 실험, (2) 역사적인 군집화(colonization) 사건, 및 (3) 화석 기록이라는 3가지 다른 분류의 자료로부터 관찰한 속도에 중심을 두었습니다. 유용한 진화 속도의 단위는 다윈이며, 이것은 한 유기체의 특성의 변화를 e/백만년 단위로 표현하는 것입니다. (여기서 e는 자연 log 의 base입니다.) 화석기록으로 관찰된 평균 속도는 0.6 다윈이며 가장 빠른 속도는 32다윈입니다. 이중 가장 빠른 속도를 나타내는 숫자는 비교를 위해서 매우 중요한 숫자입니다. 즉 현대 개체군에게서 관찰된 진화의 속도는 이것보다 같거나 커야 합니다.
야생에서 역사적인 군집화 사건을 통해서 관찰된 진화의 평균 속도는 370 다윈(필요한 속도보다 10배 정도 빠릅니다.)입니다. 사실상, 군집화 사건에서 발견된 가장 빠른 속도는 80,000 다윈으로 이는 필요한 속도 보다 2500 배나 빠릅니다. 실험실에서 관찰된 진화의 속도는 좀더 인상적으로 평균 60,000 다윈이며 빠른 것은 200,000만 다윈으로 이는 필요한 속도보다 6000배나 빨랐습니다.
좀더 최근의 논문은 야생에서 구피(역주 - 송사리과의 물고기)의 진화 속도를 4000에서 45,000 다윈의 범위에 있는 것으로 계산했습니다(Reznick 1997). 단지 400 다윈이면 마우스를 코끼리로 1만년내에 변화시키기에 충분합니다(Gingerich 1983).
빠른 진화의 가장 극단적인 사례들은 충적세의 약 10만년 동안 호미니드의 소뇌가 2배가 된 것입니다(Rightmire 1985). 이 "독특하고 놀라운" 진화의 속도는 단지 7 다윈에 지나지 않았습니다(Williams 1992, p. 132). 이 속도는 한 세대마다 아주 작은 약 0.02%의 증가로 바꾸어 말할 수 있습니다. 비교하자면, 깅그리치의 연구에서 가장 빠르게 관찰된 화석 기록은 1천년 동안 37다윈이었으며, 이것은 한 세대당 약 0.06%의 변화밖에 되지 않습니다.
반증가능성:
만약 현대에서 진화의 관찰된 속도가 화석기록에서 발견된 속도를 설명할 수 없다면, 공통조상 이론은 정당화하기가 극히 힘들 것입니다. 예를들어, 후기 신생대동안의 말의 진화 속도가 80,000 다윈보다 더 컸을 수도 있었으며 그렇다면, 현대 개체군에서 관찰된 속도로는 설명이 불가능했을 것입니다. 군집화에 의한 진화의 평균 속도가 평균 400다윈 정도이므로 만약 화석기록에서의 평균적인 속도가 4000 다윈이면 강력한 반증이 되었을 것입니다.
예측 5.8: 유전자의 변화 속도
뉴클레오타이드의 치환으로 측정한 유전자 변화 속도는 화석기록에서 관찰된 두 종간의 서열의 차이에서 허락하는 시간이 필요한 속도와 일관성이 있어야 합니다.
확인:
우리가 비교해야 하는 것은 (1) 종의 갈라지는데 필요한 시간에 대한 화석기록의 예상치 (2) 두 종간의 뉴클레오타이드의 차이 및 (3) 현대의 종들에게서 발견되는 돌연변이율이라는 서로 독립적인 자료를 비교하는 것입니다.
예를들어, 진화론적인 친척관계가 가장 잘 연구된 인간/침팬치의 분기를 생각해 보겠습니다. 침팬지와 인간은 화석기록을 근거로 볼 때 6백만년 전에 서로 갈라져 나왔거나 다른 말로 공통조상을 가지고 있습니다(Stewart and Disotell 1998). 침팬지와 인간의 게놈은 매우 비슷해서, 그들의 DNA 서열은 전체 98%가 서로 동일합니다(King and Wilson 1975; Sverdlov 2000). 이들 게놈의 가장 큰 차이는 가유전자, 비전사되는 서열, 그리고 3염기로 구성된 코돈의 마지막 염기의 변화(역주 - 4가지 아무거나 와도 아미노산에는 변화가 없는 경우가 많음)입니다. 이들 모두는 기능이나 표현형에 어떠한 영향도 주지 않기 때문에 자연 선택으로부터 자유로울 것입니다. 이들 지역은 모두 기능이 없기 때문에, 모든 돌연변이는 그들의 서열에 생기면 그대로 유지됩니다. 그러므로, 그들은 게놈의 자발적인 돌연변이의 기본 속도를 나타내는 것입니다. 가장 서열이 일치하지 않는 지역은 바로 종 간의 비교를 해야하는 곳입니다.
종이 갈라지는데 6백만년이 걸렸다면, 인간과 침팬지의 가장 서로 다른 지역의 가장 빠른 염기 치환 속도는 한 위치당 ~1.3 x 10-9 염기치환/년입니다. 한 세대가 15년에서 20년이라고 한다면 이것은 약 한 세대당 한위치에 대해서 ~2 x 10-8의 치환율과 동일한 것입니다(Crowe 1993; Futuyma 1998, p. 273).
자발적 돌연변이의 기본 속도는 암연구에서 극히 중요한 것이며, 인간에게 있어서는 매우 광범위하게 연구되었습니다. 인간의 몇몇 유전자에서 관찰된 자발적인 돌연변이율에 대한 검토에서 기본 돌연변이율은 한 세대당 한 위치에서 약 -5 x 10-8 염기치환이라는 것을 발견했습니다. 이 속도는 최소한입니다. 왜냐하면 이 값에는 유전자의 삽입, 결실 혹은 유전자를 완전히 기능을 상실하게 하는 다른 염기 치환 돌연변이를 포함시키지 않았기 때문입니다(Giannelli et al. 1999; Mohrenweiser 1994, pp. 128-129). 그러므로 세가지 독립적인 자료가 서로 잘 맞아떨어지는 것은 극히 인상적인 것입니다.
다른 종들에서도 비슷한 결과를 얻었습니다. (Kumar and Subramanian 2002; Li 1997, pp. 180-181, 191). 간단히 말해서, 관찰된 유전자의 돌연변이율은 고생물학적으로 종이 갈라지는 시간이나 혹은 게놈의 차이를 근거를 두고 추론한 값과 매우 잘 일치했습니다. 그러므로, 돌연변이에 대한 관찰된 속도는 쥐, 침팬지, 및 인간과 같은 서로 다른 종간에서 관찰되는 유전자의 차이를 잘 설명할 수 있습니다.
반증가능성
현대 유기체에서 관찰된 유전자 돌연변이율이 화석기록이나 서열의 차이를 추론해서 얻은 속도에 비해서 십의 몇승 단위로 적을 수도 있을 것이다.
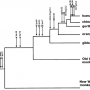 분자 서열 증거
분자 서열 증거
 중간화석에 대한 정리 -- 1
중간화석에 대한 정리 -- 1