
역주 : 이글을 읽어보면서 아마 생물학에 관심없으면 이해하지 못할 내용이 많을 뿐 아니라 해석도 하지 못할 것이라고 생각 되어서, 중간 중간에 그림을 첨가하였습니다. 이 그림은 원본에는 없는 것들입니다.
서론
Other Links:
|
다윈의 블랙박스(Darwin's Black Box,The Free Press, 1996)라는 책에서 생화학자 마이클 비히는 많은 생물학적인 시스템은 "환원불가능할 정도로 복잡"하며, 진화하기 위해서는 몇 개의 시스템이 동시에 생겨나야 한다고 주장했다. 그는 생물학에는 이러한 시스템이 존재하며, "환원불가능한 복잡성"이 존재한다는 것은 지적 설계자가 있기 때문이라고 주장했다. 비히는 매우 자세하게 생화학 시스템을 서술하고 다른 몇가지를 암시적으로 언급하면서 이들들이 "환원불가능 할 정도로 복잡하다"고 주장한다.
대부분의 일반 대중에게 초점이 맞추어진 과학 서적은 지식이 어디까지 발전했는가에 초점을 맞춘다. 즉 우리는 무엇을 알고 있는가? 이것이 무엇을 제시하는가? 그리고 우리에게 무엇이 중요한지를 말한다. 이와는 반대로 비히의 책의 특징은 무지의 경계에 대해서 보여주고 있다. 즉, 우리가 무엇을 모르는지, 지식의 무족으로 우리가 얼마나 우리 자신을 모르는지를 보여주고 있다.
실재로, 이러한 점은 비히의 책 전체의 주제이다. 그는 어떤 시스템에 대해서 보다 간단하고 작동이 가능한 형태를 생각해 낼 수 없다면 "환원불가능하게 복잡하다"고 적어 버린다. 이러한 주장을 증명할 방법은 전혀 없다. 우리가 할 수 있는 것은 사실과 논리적으로 제시된 것들을 살펴보고, 이것들이 그럴 듯한가를 판단하는 것 뿐이다. 논리가 맞는가하는 것은 완전히 다른 이야기이다. 실제로, 이 연속된 게시물은 그러한 합리적 추론이 잘못이라는 특정한 사례들을 보여주기 위함이다. 또한 몇부분은 비히가 전체적인 그림을 제시하는데 실패했기 때문에 잘못된 것이며 즉, 비히가 논리적으로 잘못된 것이라는 중요한 사실들을 우리에게 드러내지는 않았다.
비히는 쥐덫의 사례로 시작한다. 그는 표준적인 쥐덫 모형이 "환원불가능하게 복잡하다"라고 말한다. 이러한 쥐덫은 아래와 같이 구성되어 있다. (역주 : 쥐덫은 일반적으로 아래와 같이 구성되어 있다. 그림은 다른 사이트의 그림을 본인이 추가한 것임)
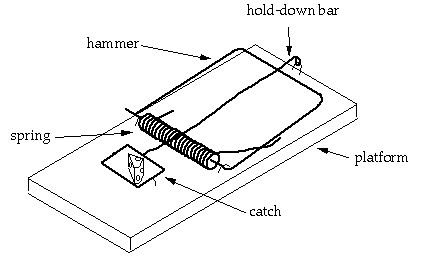
(1) 평평한 나무로 된 받침대 역할을 하는 플랫폼(platform)
(2) 작은 생쥐를 눌러서 실재로 생쥐를 잡아주는, 금속으로 된 해머 (metal hammer)
(3) 쥐덫이 작동할 때 플랫폼과 해머를 눌러주는 끝이 펴진 스프링
(4) 약간의 압력이 가해지기만 해도 민감하게 작용하는 고리 (catch) 부분
(5) 고리 부분에 연결되어 있으며 쥐덫이 동작할 때 hammer가 뒤로 튕겨서 쥐를 잡을 수 있게 해주는 금속 막대 (metal bar) (또한 전체 시스템을 잡아주는 U자 못등이 있다.)
비히는 왜 이 시스템이 "환원 불가능하게 복잡하다"고 하는지 아래와 같이 논리를 펴고 있다.
한 부분이라도 없어도 이것이 쥐를 잡을 수 있는가? 만약 나무로 된 지지대가 없다면, 다른 부품을 붙일 플랫폼이 없게 된다. 만약 해머가 없다면, 쥐는 나무로 된 지지대 위에서 잡히지 않고 밤새 춤추며 놀 수 있다. 만약 스프링이 없다면 해머와 플랫폼은 느슨하게 쨍그렁 거릴뿐이고, 이 쥐는 전혀 방해받지 않을 것이다. 만약 고리 부분이나 금속 막대가 없다면 당신이 손을 놓자마자 스프링이 해머를 움직이게 할 것이다...
만약 당신이 나에게 표준적인 형태의 쥐덫이지만 환원 불가능하지 않은 것을 제시하라고 했다고 하자. 당신은 나에게 위에 언급한 모든 부분을 준다. 나는 표준적인 것과 가장 비슷하며 한가지 부품은 사용하지 않고 실재로 작동하는 쥐덫을 만들어야 한다. 가능하겠는가?
그렇다. 나무로 된 지지대는 버려도 된다. 그러면 어디에 쥐덫을 설치하겠는가? 바닥에 설치하면 된다. 내가 쥐덫을 마루 마닥에 조립해서 설치하게 되면 뭐가 문제인가? 모든 기능을 다 하는 쥐덫이지 않은가?
물론 나는 그렇게 생각한다. 이것이 매우 유용할 것인가? 그렇지는 않다. 즉 키트 (kit)가 아닌 일반적인 쥐덫을 사는 것은 분명히 몇가지 이득이 있다. 나는 쥐덫을 조립해야 할 뿐만 아니라 돌이나, 콘크리이트 바닥 또는 바닥이 평평하지 않거나, 토양과 같이 부드러운 곳에는 이 키트를 설치할 수 없다. 이것을 기기나 가구 뒤부분이나 밑에 설치하는 것은 여간 성가신 것이 아니다.
분명히 이것은 더 열등한 것이다. 그러나 명확한 것은 이것은 사용할 수 있다는 것이다!
이것은 산뜻하게 "환원 불가능한 복잡성"의 문제를 보여준다. 이것은 주장일 뿐이다. 주장을 위해서 오직 논리와 사실들만이 사용되었을 뿐이다.
위의 글이 talk.origins에 게시되자 비히는 다음과 같이 답변했다.
이것은 매우 흥미있는 답변입니다. 하지만 당신은 당신에게 주어진 나무 지지대를 다른 것으로 교체했을 뿐입니다. 쥐덫은 지지대 없이는 아직 제대로 동작하지 못합니다.
이것은 완전히 논점을 잘못 파악한 것이다. 지지대 없는 쥐덫도 완전히 제대로 동작한다. 이것은 주변의 환경을 이용한 것 뿐이다.
비히는 계속해서 다음과 같이 말하고 있다.
게다가, 당신은 근본적으로 해체된 쥐덫을 받았으며 당신은 이것을 다시 조립했습니다. 모든 부품들은 지적인 존재에 의해서 이미 서로 적절하게 적응된 것들입니다. 내가 지적해야하는 중요한 점은, 당신이 "아무 부품도 없이" (최소한 쥐덫에 맞도록 특별히 디자인되지 않은 것으로) 어떻게 시작할 수 있는가이며, 제대로 작동하는 환원 불가능하게 복잡한 쥐덫을 어떻게 만들 수 있는가라는 점입니다.
이것은 "환원 불가능한 복잡성"의 일반적인 문제를 그대로 그러낸다. - 이것은 "간극의 신(God of the Gaps)" 설명일 뿐이다. 만약 우리가 "환원 불가능한 복잡성"이라고 제안된 것이 실제로는 그렇지 않다는 것을 보여도 지지자들은 단지 그 부분만을 제외하고 다시 새로운 시스템이 "환원 불가능한 복잡성"이라고 주장할 수 있게 된다. 이것이 제노의 파라독스와 비슷한 것은 순전히 우연이다.
Pseudogenes
지적 설계자에 대한 반대 주장의 한가지는 염색체 안의 유전자안에 포함된 엄청난 양의 허접쓰레기들이다. 인간의 유전자는 90-95%는 아마도 쓰레기이며, 쓸데 없는 서열이며, 이들중의 상당한 부분은 기능이 있는 유전자와 매우 닮아있으나, 제대로 사용하기엔 너무 낡은 것들이다(pseudogenes). Dr. Ken Miller의 pseudogene에 대한 글을 반박하기 위해서 비히는 아래와 같이 주장한다(p.226).
밀러의 주장이 설득력이 없는 두 번째의 이유는 pseudogene이 아무런 기능도 없음에도 불구하고 진화는 어떻게 이러한 pseudogene이 생겨났는지에 대해서 어떠한 설명도 하지 못한다는 것이다. 유전자의 pseudocopy를 하는 것만도 (역주 : evan을 even으로 해석했음) 수십개의 복잡한 단백질들이 필요하게 된다. 즉 DNA의 2가닥을 벌리고, 복제 기구를 정확한 위치에 배치시키고, 핵산을 짜 맞추어 나선으로 만들고, 이 pseudocopy를 다시 DNA 나선에 끼워 넣는 등등이 필요하다. 그의 논문에서 밀러는 어떻게 이러한 기능들을 하는 것들이 다윈 주의자들이 말하는 단계적인 과정을 거쳐서 일어났는지 아무것도 말하지 않았을 뿐만 아니라, 우리가 이러한 정보를 어디서 찾을 수 있는 과학 문헌도 알려주지 않았다. 그가 그할 수 없었던 것은, 이러한 정보를 어디에서도 찾을 수 없었기 때문이다.
이 문제에 대한 사실을 말하면, 그 답은 유전학교과서 어디에서나 찾을 수 있다는 것이다. 생물학에서 이러한 중복(duplication)이 일어나는 것은 2가지 기작이 있으며 이 2가지 모두 실험적으로 증명이 된 것들이다.
비히는 명백히도 엄청난 양의 한 카피의 유전자가 여러카피로 중복되는 tandem duplication에 대한 문헌을 완전히 무시하고 있다. 가장 일반적인 기작은 재조합 기구가 두 개의 염색체를 정확하게 일치시키지 못해서 일어나는 불균형 교차 (unequal crossing over) 이다. (역주 : 교차에 대한 그림 참조) 이러한 것들이 일어난다는 것을 실험실에서 보일 수 있다.
{kind=link}
예를들어, 초파리(Drosophila)의 경우 Bar와 UltraBar라고 불리는 돌연변이가 존재한다. Bar 유전자를 가진 돌연변이는 보통 구형의 눈이 매우 작고 뚜렷하게 타원형이다. Ultrabar를 가진 돌연변이는 좀더 가늘다. 만약 당신이 각각의 돌연변이 초파리의 순순하게 분리하여 키울 수 있다고 하여도 각각의 그룹에서 다른 돌연변이가 아주 낮은 빈도로 나타나게 된다. 다시 말하면, 순수한 Bar 유전자를 가진 그룹에서 가끔씩 Ultrabar 파리가 나타나며, 순수한 Ultrabar 그룹에서도 가끔씩 Bar 돌연변이 파리가 나타난다. Bar 돌연변이에서는 염색체의 한 부분이 반복(tandem duplication)되어 있다. Ultrabar 돌연변이에서는 이러한 반복이 더 많다. 세대가 지나가면서 tandem duplications가 더 많아지거나 (Bar 돌연변이가 Ultrabar로) 반복된 경우가 많은 경우는 수가 줄어드는 것(Ultrabar 돌연변이가 Bar 돌연변이로)되는 것은 세포의 재조합 기구에 의한 피할 수 없는 결과이다.
두 번째 기작은 역전사(reverse transcription)와 integration이다. 이 경우에는 유전자의 mRNA가 DNA 조각으로 역전사 되며, 이것은 다시 유전자 안에 끼워넣어진다. 역전사 효소는 언제나 사용이 가능하며 이것은 세포안의 많은 retrotransposons과 결함을 가진 레트로 바이러스들 때문이다. Integration 단계는 homologous recombination 혹은 다른 무작위적인 과정에 의해서 일어난다. DNA 조각이 세포에 넣어지면 유전자 안에 끼워넣어지는 경우는 매우 낮은 빈도로 일어나며, 많은 유전공학 기술은 이것에 의존한다. 많은 pseudogene들과 일부 기능을 가지고 있는 유전자는 이러한 방식으로 만들어졌다는 분명한 흔적을 가지고 있다. 즉 이들은 부모 유전자에 비해서 인트론이 없고 긴 뉴클레오타이드 A를 유전자 뒤에 가지고 있다. (신핵세포의 mRNA는 마지막 부분에 A가 길 게 늘어져 있다. (역주 : DNA의 염기중에 A를 말하는 것입니다. 흔히 poly A tail이라고 부릅니다.)
그러므로 우리는 생물학적인 지식으로 볼 때 시간이 충분하다면, pseudogenes은 불가피한 현상이라고 예측할 수 있다. 비히가 pseudogene 형성에 필요하다고 주장한 복잡한 기구(machinery)는 모든 살아있는 계 (system)에 존재할 뿐만 아니라, 완전히 다른 목적 때문에 존재한다.
진화는 pseudogenes이 생겨날 것이고 서서히 없어지거나 혹은 완전히 제거될 것으로 예측하며, 공통 조상은 공통의 pseudogene을 공유할 것이라고 예측한다. 진화는 또한 실제 유전자에서 pseudogenes을 구분하는 방법을 제공한다. (별 변화를 주지 않는 돌연변이와 그렇지 않은 돌연변이를 조사함으로써)
Cascades
분자 생물학 분야에서 중요한 주제중의 하나가 유전적인 cascade 으로 하나의 유전자가 다른 유전자를 촉발시키고, 촉발된 유전자가 다른 유전자를 촉발시크는 것이다. 비히가 인용한 cascade는 2가지가 있는데 혈액응고와 보체계(complement system)이다. 보체계는 complement system는 항체에 의해서 활성화되는 여러 가지 단백질로 이 단백질들은 침입한 박테리아의 막에 구멍을 내서 박테리아가 살아나가기 위해 필요한 전해질과 이온의 균형을 파괴한다. (역주 보체계는 보통 생화학 교과서 보다는 면역학에서 다루게 됩니다. 인터넷에서 1, 2, 3 등에서 내용을 찾아볼 수 있다.) 비히의 주요 주장은 생화학적인 연쇄반응은 "환원불가능한 복잡성"을 보인다는 것이다. 이 주장은 이 연쇄반응에서 모든 구성요소중 하나라도 없으면 전체 연쇄반응이 기능하지 않게 되면 그렇기 때문에 알려진 진화적인 과정으로는 연속적인 반응단계의 추가를 가지고는 이러한 연쇄반응을 만들어낼 수 없다는 것이다. p.87:
연쇄반응의 본질상, 새로운 단백질은 즉시 조절되어야만 한다. 새로운 단계가 연쇄반응에 포함되는 처음부터 proenzyme이 필요하고 적절한 시기에 proenzyme을 반응하게 하는 활성화 효소가 필요하다. 각 단계가 여러 가지 구성요소로 되어 있기 때문에 혈액 응고 계의 전체가 환원불가능한 복잡성을 보일 뿐만 아니라 각 단계도 마찬가지이다.
우리는 쉽게 위의 문장의 대부분이 거짓임을 보일 수 있다. 이것은 진화론적인 과정으로 가정할 수 있다. 게다가, 우리는 이러한 과정에서부터 이러한 과정에서 예상되는 결과로 나아갈 수 있으며, 그러므로 생명계에서 어떠한 것이 발견될지를 예측할 수 있다.
실제로, 이 기작의 힌트가 비히의 책에서 발견된다. 분명히 연쇄반응은 시작이 있어야 한다. 어떻게 시작할 수 있는가? p.83:
... 혈류중에 트롬빈의 양은 매우 적다. 혈액응고에서 연쇄반응에 관여하는 단백질들은 같은 단백질이 더 많이 생기도록 촉진시키기 때문에 그러므로 자기 촉매반응이다.
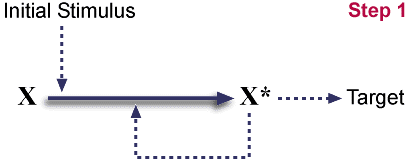
Starting pathway.
X<---X* 과정은 아래 도식에서 생략되었다. 이것은 자발적인 과정이거나 혹은 X*가 촉매가 되는 반응이다. (이 두가지 사례가 모두 존재한다는 것이 알려졌다.) 자극을 가하는 촉발이 일어나면 X는 X*가 바뀌게 되며 X*는 자기 촉매반응으로 이러한 반응을 촉진시킨다. X*는 target에 작용한다.
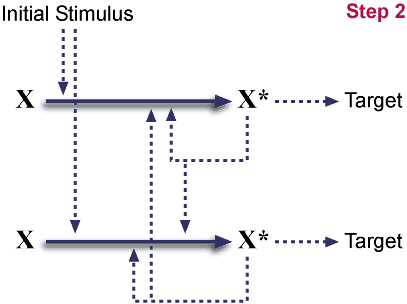
Post gene duplication.
이것은 완전히 대칭적인 계이다. (도식에서는 대칭적이지는 않지만, 하나로 배치될 수 있다.) 이러한 배치는 중성적이다. 즉 각 화학종들은 어떠한 이득도 없다. 한편 이러한 중복은 매우 흔한 사건이다.
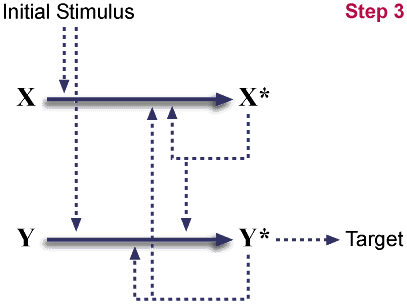
Mutation eliminates X*'s ability to interact with target.
중복된 것은 이제 좀더 알아보기 쉽게 하기 위해서 Y/Y* 라고 표기했다. 다시 말하지만, 이들 유전형은 중성적이다. 즉 더 이롭지도 않고 더 나쁘지도 않다. (역주 : 돌연변이가 일어나는 경우에도 활성에 전혀 영향을 주지 않는 경우가 많다.) 이러한 중복이 일어나는 경우 돌연변이가 일어나서 이중 하나가 활성이 없어지는 경우는 매우 흔하다. (돌연변이가 일어나서 한 복사분을 완전히 불활성화시키는 것은 매우 흔할 것이며, 이는 시스템을 원래의 상태로 만들어 놓는다.)
돌연변이가 몇군데 일어나는 것이 이제 가능해진다:
- X 는 돌연변이에 의해서 비활성화 된다.
- 시스템은 처음으로 돌아가게 된다.
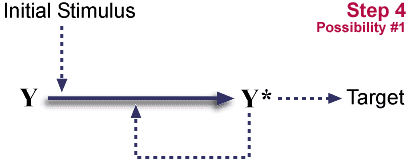
- Y* 의 X->X* 와 Y->Y* 촉매 반응을 일으키지 못하게 된다.
시스템은 이제 X와 Y에 의존하게 된다. (자극이 한가지인 경우 유용한 활성화에는 충분하지 않게 된다.) 이것은 "환원 불가능한 복잡"하게 된다.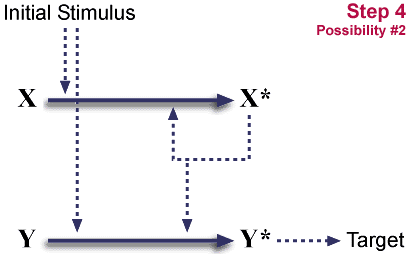
- Y의 자극에 대한 반응하는 능력이 소실된다.
시스템은 이제 X 와Y에 의존하게 된다. 이것은 "환원 불가능한 복잡"하게 된다.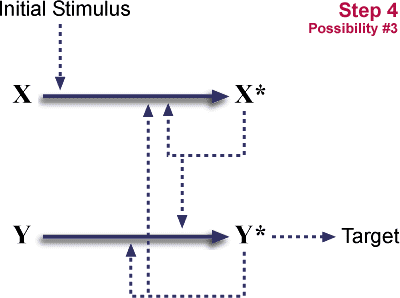
- 시스템은 처음으로 돌아가게 된다.
그러므로 중복과 몇몇 기능의 소실, 그리고 기능을 소실하게 된 2개의 돌연변이중 한가지는 한 단계로 과정을 2단계의 연쇄반응으로 만들었다. 처음 단계는 중성적으로, 더 이익이 되지도 더 불리한 것도 아니다. 이러한 중성적인 돌연변이는 주기적으로 일어난다. 마지막 단계는 연쇄반응에서 고정되어 있다. 이것은 아마 이익이 많을 것이다. 왜냐하면 연쇄반응이 다단계로 이루어진 경우 보다 조절이 용이하며 (장기적인 이익) 증폭 (하나의 X*가 여러개의 Y를 활성화시키면서 신호를 증폭시킨다.)에 유리하기 때문이다.
위의 2단계의 과정은 각각 다시 2단계씩으로 나뉘어질 수 있는 것은 명백하며, 이러한 기작으로 인하여 여러단계의 연쇄 반응으로 확장될 수 있다. 그리고, 다시 만들어진 2단계는 모두 자기 촉매 반능을 가질 수 있으며 또 새로운 단계로 확장될 수 있다. 이 얼마나 많은 "환원 불가능한 복잡성"인가.
이것은 물론, 하나의 모델이다. 모델은 예측을 할 수 있게 해주어야 하며 이 모델에서도 예측이 가능하다. 만약 반응 경로가 이러한 방식으로 진화했다면, 반응 경로중의 서로 연속적인 단계는 서로 서열이 비슷한 단백질에 의해서 촉매될 것이다. (비히가 제시한 2가지 사례인, 혈액 응고와 보체계 연쇄 반응의 한가지 특징) 만약 단백질들이 인접한 유전자(tandem duplication)에 의해서 염기서열이 보존된다거나, 혹은 역전사된 명백한 흔적 (인트론이 없거나 혹은 poly-A 꼬리가 genomic sequence에 포함되어 있거나) 또는 둘다면 더 좋다. 우리는 또한 좀더 적거나 혹은 더 많은 단계를 가진 연쇄반응 시스템을 발견할 가능성도 있다.
이것이 보체계와 혈액응고 연쇄반응에 해당할까? 고백한다면, 나는 이것을 명확하게 알지 못한다. 비록 대단한 검사 방법은 아니지만, 이 두가지 시스템에서 많은 단백질의 서열이 매우 비슷하다는 기준은 만족시킨다. 좀더 강력한 검증 그리고 "환원 불가능한 복잡성" 가정에 대한 위반사례는, 이러한 연쇄반응계의 변형이 생명계에서 발견되는 것이다. 예를들어 만약, 내가 인간의 것보다 한단계 적은 시스템을 찾아낸다면 이것은 인간의 시스템이 환원 불가능한 복잡한 것이 아니게 되며 비히의 "전체 혈액 응고 시스템이 환원 불가능한 복잡성을 보일 뿐만 아니라 각 단계도 마찬가지이다"라는 주장을 반박하는 것이다. 다시 말하지만, 이것은 나의 전문분야가 아니며 나는 이것에 대해서 잘 모른다. 사실상, 이 분야는 탐험되지 않은 분야로 남아있다. 어류에서도 인간과 동일한 방식으로 혈액이 응고되는가? 무악어 (먹장어와 칠성장어와 같은 턱뼈없는 물고기 - 무악어류)는? lancets(척추의 원시적인 형태 (역주 - 척색을 말함)을 가지고 있는 무척추 동물) 은 어떠한가 ? (역주 : lancet은 창고기(lancelet) 를 잘못쓴 것이 아닌가 싶음. 창고기는 성체에 척색이 있음) 비히는 이러한 점에 침묵하고 있으며, 그가 반대 모델에 관심조차 없는 사실은 혹은 더 나쁘게, 그가 그의 모델로부터 예측되는 것은 전혀 고려하지 않는다는 것은 좋은 징조는 아니다. 실재로 자신의 모델을 비평적인 시각으로 살펴보는 것은 과학과 화물 숭배의 차이이다. (역주 : 화물 숭배는 간단히 말하면 화물 자체를 신으로 숭배하는 것인데 자세히 알고 싶다면 마빈 해리스의 "문화의 수수께끼" 와 같은 책을 보면됩니다. )
The Krebs Cycle 크렙스 회로
크렙스 회로 (또는 구연산 회로 혹은 시트르산 회로로 알려짐, 역주 : 보통 국내에서는 TCA 회로라고 합니다. 그림 참조)는 대사에 중요한 부분이다. 대부분의 생물학 교실은 이것을 최소한 약간이라도 다루며, 생화학 교실에서는 매우 자세하게 분석한다. (나는 이것을 암기해야만 했었다.)
{kind=link}
크렙스 회로는 대사의 다양한 기능의 중심이다. 포도당 분해의 2번째 단계이며, 다른 연료를 태우는데 사용되기도하며, 다양한 탄소 유닛을 만드는데 유용하는 등 여러 가지 기능이 있다. 9개의 효소가 크렙스 회로를 돌린다. 언 듯 보기엔 이것은 세포의 가장 중요한 근본원리로 보이며 "환원 불가능한 복잡성"을 보인다. 어떤 효소도 제거하면 분명히 회로를 더 이상 돌아가지 않을 것이다.
불행히도, 비히는 그의 책에서 크렙스를 언급하지 않았다. 이것은 복잡한 생화학적인 시스템이며, 분명히 그의 논문이 매달릴 만한 훌륭한 고리가 될 것이다. 아닌가?
그러나, 문헌을 자세히 살펴보면, 크렙스 회로의 "환원 불가능한 복잡성"가설과 같은 것의 문제점이 발견된다. 일부 시스템에서는 glyoxylate shunt 라는 pathway가 존재하며 이것은 회로를 가로질러 진행한다. (역주 : glyoxylate shunt 회로의 그림 참조. 이 그림에서 glyoxylate 회로를 이용하면 fumarate로 진행하지 않는다.) 그리고, 다른 변형된 회로들이 존재한다. 그러므로 크렙스 회로는 가장 최소한의 돌에 새겨진 회로가 아니라, 여러 가지 대체 회로들이 존재한다. 게다가, 일부 종들은 크렙스 회로가 완전히 없어도 생존할 수 있다. 예를들어 박테리아 Haemophilus influenzae 의 모든 완전한 DNA 염기 서열이 결정되었는데 이것은 크렙스 회로의 몇몇 단계의 유전자가 완전히 없다는 것을 확인시켜 주었다.
{kind=link}
이것은 의문이 생기게 한다. 만약 크렙스 회로의 복잡함도 "환원 불가능한 복잡성"이 아니라면, 도대체 "환원 불가능한 시스템"을 인식하기 위한 우리의 능력을 어떻게 신뢰할 수 있는가? 결국, 우리가 이것을 인식하기 위한 유일한 기준은 바로 합리적인 진화론적인 단계를 추론할 수 없을 때 뿐이다.
비히의 주장의 한가지는 과학적인 논문중에서는 복잡한 생화학적인 계의 진화에 대해서 그럴 듯한 다위니즘의 모델을 자세하게 묘사한 것이 없다는 것이다. p.179:
복잡한 생화학적인 계의 진화에 대해서 자세하게 다룬 미팅이나, 책, 혹은 논문 같은 것은 전혀 없었다.
나는 그가 그의 다음번 책/판에서는 이것을 고쳤으면 하고 바란다. 왜냐하면 이것은 완전히 거짓이기 때문이다.
"다윈의 블랙박스"가 출판되는 것과 동시에 다음과 같은 논문이 나왔다.
The puzzle of the Krebs citric acid cycle: Assembling the pieces of chemically feasible reactions, and opportunism in the design of metabolic pathways during evolution.
Journal of Molecular Evolution, Sep 1996, 43: 293-303
Melendez-Hevia, Waddell & Cascante
이들 저자들은 비히의 주장에 의하면 지금까지 완전히 무시된 바로 그 문제들을 다루고 있다. 즉 어떻게 복잡한 생화학적인 pathway가 다윈 주의자들이 말하는 과정을 통해서 그리고 각 단계는 선택될 수 있는 이점을 유지하면서 진화될 수 있었는지를 다루고 있다.
비히에게 더욱 않된 것은 이 논문의 관계논문을 찾아보면 다른 생화학적인 경로에 대해서 분석한 것들이 여러번 인용되었다는 것을 알 수 있다. 예를들어, 같은 저자들이 5탄당 경로의 진화에 대해서 1985, 1988, 1990, 그리고 1994년까지 4번이나 논문을 제출했으며 포도당 생합성에 대해서 1993년부터 논문을 제출했다. 그들은 크렙스 회로의 진화에 대해서 1981 (x2), 1985, 1987 (x2), 1992 논문들과 1992년 이후의 것으로 대사 진화의 일반적인 주제에 대한 책들도 인용하고 있다.
비히의 문헌 조사는 1994년까지 아무것도 찾아지지 않았다고 한다.
이것에 용기을 입어, 나는 아미노산 생홥성에 대한 논문을 Entrez 에서 간단히 찾아봤다(가물 거리는 기억을 되살려서). 충분히 많이 나왔으며, 아래의 몇가지를 포함하여 한 뭉치의 논문을 찾는 것은 문제도 되지 않았다.
- Orig Life Evol Biosph 18: 41-57 (1988)[88217276]. New prospects for deducing the evolutionary history of metabolic pathways in prokaryotes: aromatic biosynthesis as a case-in-point. S. Ahmad & R. A. Jensen
- Mol Biol Evol 2: 92-108 (1985)[88216112]. Biochemical pathways in prokaryotes can be traced backward through evolutionary time. R. A. Jensen
- Microbiol Sci 4: 258, 260-2 (1987)[91058939]. Enzyme specialization during the evolution of amino acid biosynthetic pathways. C. Parsot, I. Saint-Girons & G. N. Cohen
- Annu Rev Microbiol 30: 409-25 (1976)[77043263]. Enzyme recruitment in evolution of new function. R. A. Jensen
- Proc Natl Acad Sci U S A 76: 3996-4000 (1979)[80035004]. Origins of metabolic diversity: evolutionary divergence by sequence repetition. L. N. Ornston & W. K. Yeh
그러므로 우리는 이제 한가지 질문을 하게 된다: 만약 효소 반응의 매우 복잡한 회로인, 크렙스 회로가 "환원불가능한 복잡성"을 보이지 않는다면, 우리는 어떻게 다른 시스템이 환원 불가능하게 복잡하다는 것을 알 수 있을까?
우리는 다음과 같은 결론을 내리게 된다 : 만약 비히가 그의 문헌 조사에 대해서 저널에 제출한다면 어떠한 것이 가장 좋을까? 아마도 "재현성 없는 결과들의 연대기"일 것이다.
What Good is An Antibody?
비히는 그의 책의 제 6장을 면역계의 일부에 할애하였다. 항체 생산에 대한 것을 다루었는데 그는 이것이 "환원 불가능한 복잡"한 과정이며 그러므로 이것은 진화의 산물이 아니라고 주장했다. 그는 아래와 같이 흥미있는 내용을 썼다.
항체들은 장난감 다트와 비슷하다. 즉 이들은 어떤 것에도 해가되는 것이 아니다. 흉가의 저주의 사인과 마찬가지로 혹은 제거될 나무의 오렌지 색의 X 자의 페인트와 마찬가지로 항체는 단지 다른 시스템에게 표시된 물체를 제거하라는 신호에 불과하다.
비히는 그런 다음에 보체계에 대해서 설명하고 있다. 그리고 다른 것은 없다. 마치 보체와 항체가 모두 환원 불가능한 복잡성을 보이며, 각각 기능하기 위해서는 서로가 필요하다는 인상을 남기고 있다. (역주 : 비히의 책을 읽어보지 않아서 잘 모르겠지만 비히가 언급한 것은 보체계의 고전적 경로 "classical pathway"인 것 같습니다. 일반적으로 보체계는 선천적인 것이지만 이 과정중에 미생물을 코팅하는 경우 후천적으로 항체가 코팅되는 경우에도 활성화 됩니다. 이글의 저자는 보체계가 항체가 꼭 있어야만 하는 것이 아니라는 것을 말하는 것입니다. 항체가 보체계를 활성화시킬 수 있지만 필수 불가결한 것은 아닙니다.)
생물학적인 진실은 이것과는 상당히 다르다. 항체는 보체가 전혀 없어도 작용할 수 있다. 실제로 비히가 틀렸기 때문에 매년 수천명의 어린이들이 살아남는 것이다.
항체는 다른 물체에 결합한다. 독을 가지고 있다고 있다고 하자. 이 독은 인체의 특이적인 기작을 이용해서 어떤 분자에 결합한다. 독의 특정한 원자는 결합하는 분자의 특정한 원자와 상호반응한다. 만약 항체가 여기에 결합하게 되어 독의 이들 원자를 뒤짚어 씌우면 독은 더 이상 독으로서 기능을 가지지 못한다. 이것이 바로 DPT 백신이 효과가 있는 이유이다. 이 백신은 독에 결합하는 항체를 만들어서 독을 중화한다.
비슷한 방식으로 많은 바이러스들이 세포에 감염될 때, 특정 분자에 의존하며 바이러스들은 이러한 목적을 하는 작은 분자 힌지 (hinge)를 가지고 있다. 만약 항체가 이 힌지(hinge)에 결합하고 끼어들 수 있다면 바이러스는 기능을 잃어 버리게 된다. 마치 문과 문틀 사이에 쐐기를 끼워 놓아서 문이 움직이지 않는 것과 같다.
항체가 단지 장난감 다트나 단지 마커보다 더 나은 다른 방식의 반응도 있다. 항체는 대칭적이다. 하체는 양쪽으로 똑 같은 팔을 가지고 있다. 그러므로 항체는 중매장이처럼 행동할 수 있다. 즉 2개의 외래물질을 서로 묶을 수 있으며 이렇게 되면 농도가 높아지게 되고 좋은 목표가 된다. 만약 침입자의 항체의 목표부분이 반복되어 있다면(바이러스의 반복적인 구조 혹은 박테리아에 매우 흔한 단백질 처럼) 항체들은 더 효과적으로 서로를 묶어줄 수 있다. 이렇게 서로 묶여 버리면, 목표단백질은 용액에서 침전된다. (역주 : 항체가 일으키는 여러 가지 반응이 있는데 이 경우는 침강반응이라고 합니다. )
나는 항체 각자가 2개의 동일한 팔을 하지고 2곳의 목표에 결합할 수 있다고 했다. 이것은 정확한 표현은 아니다. 항체중에는 IgM이라고 하는 것이 있으며 이것은 5개의 분자로 이루어져 있어서 10개의 목표 분자와 결합할 수 있다. (역주 : 일반적으로 5개의 분자가 결합되어 있지만 6개 짜리도 인간과 생쥐에게서 발견됩니다.) 이러한 배치 때문에 IgM은 깊숙히 파묻혀 있는 목표를 서로 연결시키기도 한다.
침입한 물질이 이러한 방식으로 응집이 일어나면, 식세포의 더 손쉬운 목표가 된다. 식세포는 또 다른 방어라인으로 항체가 없어도 기능을 할 수도 있다.
항체는 무엇인가? 어디서 유래한 것인가?
내가 믿기로는 이것은 아직 풀리지 않은 문제이다. 하지만 이번에도, 비히는 이것에 대해서 흥미있는 힌트를 남기고 있다. 이 힌트중에는 비히의 주장을 부정하게 하는 것이 있다.
예를들어, 비히는 포유류에서 항체를 생산하는 시스템에 대한 많은 주장을 하고 있으며 이것들이 얼마나 "환원 불가능한 복잡성"을 보이는가를 주장하고 있다. (Chapter 6; especially p.130-131). 가장 주된 주장의 하나는 아래와 같다.
- 이 스스템은 수천, 수만가지의 독특한 항체를 만들지만 각각은 특이성이 다 다르다. 이 시스템에서 생산되는 것중에서 오직 몇 개 혹은 오직 한 개정도만이 쓸모 없는 것이다.
- 이 시스템은 항체유전자를 짤라내는 생화학적인 기구(machinery)가 없으면 작동하지 않는다. 이제 다시, 이 복잡한 기구는 항체의 유전자가 없으면 아무런 기능도 없다.
Scientific American 1996년 11월 호에는 면역계의 진화에 대한 2개의 논문이 실렸다. 흥미롭게도 이들 논문은 비히의 주장을 모두 반박하는 증거를 제공한다.
첫째로, 일부 나방에서는 헤모린(hemolin)이라는 단백질을 생산하며 이 단백질은 박테리아에 결합하여 곤충의 hemolymph라고 하는 혈액과 유사한 물질에서 박테리아를 제거하는데 도움을 준다. 헤모린의 단백질의 서열은 척추동물의 항체와 관련이 있다는 것이 드러낸다. 단 하나의 항체는 쓸모없다는 가정에 대해서 많은 것을 제공한다.
두 번째 논문은 상어의 항체에 대해서 설명하고 있다. 인간에 있어서 항체의 유전자는 서로 조합이 일어나서 다양한 DNA 조각을 만들며 이것은 염색체에 줄지어서 있다. 각각의 면역계의 세포에서 항체 유전자는 아래의 방식대로 재조합된다.
| Genome: | V1-V2-V3-V4-D1-D2-D3-J1-J2-J3-C |
| Possible Antibodies: | V1-D3-J1-C |
C 는 항체의 불변 영역이며, V,D,J 조각들은 (Variable, Diversity, and Joining) 아주 많은 조각들의 집단에서 유래한 것이다. (역주 : 좀더 자세한 그림)
{kind=link}
그러나, 상어의 경우 genome에서의 배치는 아래와 같다.
| V5-D5-J5-C | V6-D6-J6-C | V7-D7-J7-C |
(Note: 여기서 숫자는 단지 표지일 뿐이며 별 의미가 없는 것임)
즉 상어의 경우에는 이미 유전자가 조립되어 있으며 이들 유전자들은 조립된 유전자의 긴 줄에 배치(tandem arrays)되어 있다. 분자 유전학적 과정을 통해서 이러한 tandem array(매우 유사한 DNA가 인접해서 배열되는 것)를 만드는 것은 잘 알려져 있다. 또한 한단계의 같은 과정을 통해서 상어의 항체 유전자 다음과 같이 만들 수 있다. (이것은 인간의 경우와 조금 비슷하게 보인다)
| V5-V6-D6-J6-C | V7-D7-J7-C |
더욱 tandem 중복에 의해서 유전자가 늘어나면서 일부가 결실된다면 인간에서 볼 수 있는 것 같은 배열이 쉽게 만들어질 것이다. 물론 이 배열은 splicing 이 되지 않는다면 별로 유용하지 않다. 하지만 상어의 예는 분명히 항체의 다양성을 위해서는 스플라이싱(splicing machinery)이 필요하다는 비히의 주장이 틀렸다고 증명하고 있다.
진화론적으로 볼 때 어떻게 시작해서 이 매력적인 시스템으로 되었는가에 대한 여러 가지 힌트가 있지만 이러한 것은 비히의 책에는 언급되어 있지 않다. 예를들어, 항체의 분자 구조는 면역 인식 (T-cell receptors)와 Cell-to-Cell 인식을 하는 분자들과 관련이 있는 구조를 하고 있다. 실재로, 항체를 만드는 재조합 기구가 왜 비 면역 세포들 예를들어 뇌의 커다란 부분을 차지하고 있는 가를 아직 신비로 남아있다. 항체가 cell-to-cell 인식 분자에서 진화한 것일까? 우리는 알 수 없다. 하지만 흥미를 끄는 이야기이다.
포유류의 면역 체계의 특징중 다른 척추동물에서는 보이지 않는 것이 무엇일까? 우리는 이미 상어의 면역 체계가 거의 비슷은 하지만 분명히 다르다는 것을 보았다. 척추를 가지고 않은 척색동물은 어떠한 면역 체계를 가지고 있을까? 나는 별로 아는 것이 없다. 하지만 그것은 분명히 면역 기능에 대한 흥미있는 어떤 것을 우리에게 말해줄 것이다. 그리고 인간에게는 있지만 그 시스템에는 없는 것은 무엇이든지 분명히 "환원 불가능한 복잡성" 주장을 손상시킬 것이다.
초벌 번역일 : 2001 5월 26일
번역을 마치고 :
창조론자들의 글은 사실 번역이 무척 어렵습니다. 왜냐하면, 일반적인 글이라면 교과서를 찾아보면서 생각하면 굳이 단어 몇 개 몰라도 되지만, 창조과학자들은 교과서를 자기 맘대로 해석하기 때문에 도움이 되지 않죠.
번역은 초벌이고 일단 올려 드립니다. 제가 모든 내용을 알고 있는 것이 아니라서 틀린 번역이 있을 수도 있습니다.