
1차 직역이고 본인은 이쪽과 고생물학이나 지질학 전공이 아니므로 수 많은 오역이 있을 것이므로 다른 곳으로 옮기는 것을 허락할 수 없습니다. 전공하신 분들이 틀린 것을 많이 수정해 주셨으면 합니다.
양서류에서amniotes(최초의 파충류)까지 전이
고대, 커다란 양서류와 최초의 파충류 사이의 가장 큰 차이점은 양막이 있는 알이다. 다른 차이는 강한 다리와 girdle과 다른 척추와 강한 턱 근육등이면 자세한 것은 Carroll (1988) and Gauthier et al. (in Benton, 1988) 을 보기 바란다. amniotes는 껍질이 있는 알을 낳는다. 이렇게 됨으로써 그들은 굳이 올챙이 시절을 거치지 않고 살아갈 수 있게 된다.
- Proterogyrinus 또는 초기의 anthracosaur (late Mississippian) -- 고전적인 labyrinthodont-양서류 두개골과 치아를 가지고 있으나, 척추, 상완골, 발까락은 파충류의 것이다. 양서류의 발목형태이다. 아직도 어류의 두개골 힌지를 가지고 있다. 5개의 발까락을 가진 앞발과 발까락의 뼈는 2-3-4-5-3 (대부분의 파충류)개로 이루어졌다.

- Limnoscelis, Tseajaia (late Carboniferous) -- 초기의 anthracosaurs에서 분화한 양서류, 하지만 파충류의 모습을 띠고 있다. braincase의 구조, 파충류를 닮은 턱 근육, 확장된 neural arches(신경궁)를 가진다.
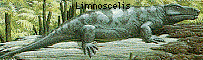
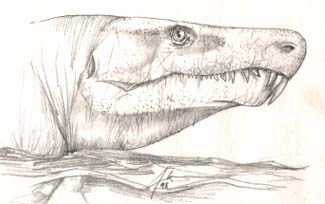
- Solenodonsaurus (mid-Pennsylvanian) -- 완전하지 못한 화석이다. 아마도anthracosaurs 와 고두류( 固頭類,cotylosaurs)의 중간일 것이다. 구개의 독아(fang)가 없어짐, 머리의 옆줄이 없어짐., Still just a single sacral vertebra, though.
- Hylonomus, Paleothyris (early Pennsylvanian) --
 이들은protorothyrids이며, 매오 초기의 고두류( cotylosaurs, 원시 파충류)이다. 매우 작고, 도마뱀만한 동물이며 양서류비슷한 두개골을 하고 있다 (amphibian pineal opening, dermal bone, etc.),어깨, 치골, 사지, 중간형태의 치아와 척추는 양서류를 닮았으며, 나머지 뼈는 파충류를 닮았으며, 파충류형의 턱근육, palatal fangs이 없고, spool-shaped vertebral centra를 가지고 있다. 아직 고막은 없을 것같다. 이러한 파충류에서 나타나는 새로운 특징의 상당수는 작은 양서류에서 보이기 때문에 (이들은 또한 가끔육지에서 부화하는 알을 낳는다) 이러한 특징은 아마도 작은 크기의 파충류에게서 처음 나타났을 것으로 보인다.
이들은protorothyrids이며, 매오 초기의 고두류( cotylosaurs, 원시 파충류)이다. 매우 작고, 도마뱀만한 동물이며 양서류비슷한 두개골을 하고 있다 (amphibian pineal opening, dermal bone, etc.),어깨, 치골, 사지, 중간형태의 치아와 척추는 양서류를 닮았으며, 나머지 뼈는 파충류를 닮았으며, 파충류형의 턱근육, palatal fangs이 없고, spool-shaped vertebral centra를 가지고 있다. 아직 고막은 없을 것같다. 이러한 파충류에서 나타나는 새로운 특징의 상당수는 작은 양서류에서 보이기 때문에 (이들은 또한 가끔육지에서 부화하는 알을 낳는다) 이러한 특징은 아마도 작은 크기의 파충류에게서 처음 나타났을 것으로 보인다.
이 조상 양서류는 약간 약한 두개골을 가지고 쌍을 이룬 "aortas" (systemic arches)를 가지고 있다. 최초의 파충류는 이 특징을 두가지 방식으로 변화시키면서 빠르게 나누어졌다. 한 계보는 한 개의 aorta를 오른쪽으로 발달시키고,방골(方骨, uqadrate)을 아래와 앞으로 swinging 하여 두개골을 튼튼하게 하였고 결과적으로 커다란 otic notch (그리고 나중에 특별히 더 변경하지 않고 청각을 발전시킬 수 있게 되었다.)를 초래했다.  이 그룹은 3개의 그룹으로 다시 나누어졌는데 그것은 두개골의 옆부분의 구멍인 'fenestrae'의 갯수에 의해 구분되는데. 무궁류anapsids (구멍이 없음), 이들에서 거북이가 나왔다; 이궁류(구멍이 2개), 여기에서 공룡과 조류가 나왔으며, offshoot 그룹은 the eurapsids (2개의 구멍이 하나로 합쳐짐)에서 ichthyosaurs가 나왔다.
이 그룹은 3개의 그룹으로 다시 나누어졌는데 그것은 두개골의 옆부분의 구멍인 'fenestrae'의 갯수에 의해 구분되는데. 무궁류anapsids (구멍이 없음), 이들에서 거북이가 나왔다; 이궁류(구멍이 2개), 여기에서 공룡과 조류가 나왔으며, offshoot 그룹은 the eurapsids (2개의 구멍이 하나로 합쳐짐)에서 ichthyosaurs가 나왔다.
파충류의 다른 주요 계보는 aorta를 왼쪽의 것만 발달시키고, quadrate bone을 위와 뒤쪽으로 이동하여 강하게 하였으며 otic notch의 필요성이 없어졌다 (청력의 향상을 위해서는 턱이 관계하는 것이 필수가 되어버림) 이들은 한 개의 구멍을 발달시켰으며, 이 그룹은 단궁형 파충류이다. 이들은 다른 파충류와는 매우 다른 길을 가게 되는데, 항온, 커다란 두뇌, 좋은 청력과 효과적인 치아를 가지게 된다. 이들중의 한 그룹은 therapsid라고 부르는데 이들은 이러한 변화가 매우 커서 결국 포유류가 되었다.
파충류 사이의 몇몇 전이
파충류는 매우 종류가 많기 때문에 몇가지만 다루어 보겠다. .. 초기 파충류에서 거북이까지만 다루어 본다.: (Also see Gaffney & Meylan, in Benton 1988)
- Captorhinus (early-mid Permain) -- protorothryids에서 바로 진화한 종류이다.
 여기에 초기 무궁형 파충류에 대한 논란의 여지가 있는데 양쪽이 모두 captorhinids에서 유래했는데 이들이 거북이의 조상이되었을 것이다. Reisz & Laurin (1991, 1993)는 거북이가 거북이를 닮은 두개골을 가진 procolophonids(페름기 말기)에서 유래했다고 믿고 있으며 다른쪽은 특히 Lee (1993) 는 거북이의 조상이 pareiasaurs라고 생각한다:
여기에 초기 무궁형 파충류에 대한 논란의 여지가 있는데 양쪽이 모두 captorhinids에서 유래했는데 이들이 거북이의 조상이되었을 것이다. Reisz & Laurin (1991, 1993)는 거북이가 거북이를 닮은 두개골을 가진 procolophonids(페름기 말기)에서 유래했다고 믿고 있으며 다른쪽은 특히 Lee (1993) 는 거북이의 조상이 pareiasaurs라고 생각한다:
- Scutosaurus 또는 다른 pareiasaurs (페름기 중기) -- 덩치가 크고 뚱뚱한 초식 파충류이며 거북이같은 뼈모양을 하고 있다. 몇몇 과는 피부에 뼈로된 판이있으며, 아마도 이것이 거북이 등껍질의 최초의 모양이 아닌가 한다.
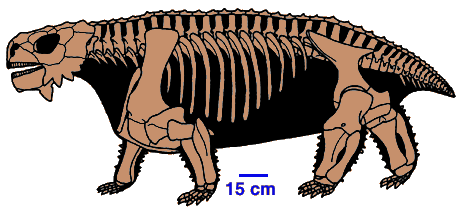
- Deltavjatia vjatkensis (Permian) -- 최근에 발견된 pareiasaur로 여러 가지 거북이를 닮은 특징 즉 두개골 (e.g., a very high palate), 사지, girdles, 그리고 마치 거북이 등가 같은 방식으로 척추의 일부가 옆으로 나왔다. (Lee, 1993)
- Proganochelys (late Triassic) -- 원시적인 거북이, 완전한 거북이같은 두개골,
 주둥이, 등껍데기를 가지고 있다. 그러나 원시적인 특징 예를들면 구강에 작은 이빨이 줄지어있으며, 아직도 확인할 수 있는 쇄골이 있으며, 단순하고 captorhinid타입의 턱 근육구조, 원시적인 captorhinid-타입의 귀, 뒤로 뺄 수 없는 목등의 특징을 가진다..
주둥이, 등껍데기를 가지고 있다. 그러나 원시적인 특징 예를들면 구강에 작은 이빨이 줄지어있으며, 아직도 확인할 수 있는 쇄골이 있으며, 단순하고 captorhinid타입의 턱 근육구조, 원시적인 captorhinid-타입의 귀, 뒤로 뺄 수 없는 목등의 특징을 가진다..
- 최근에 발견된 쥬라기의 거북이는 아직 자세하게 알려지지 않았다.
쥬라기 중기의 거북이들은 벌써 2가지로 나뉘어진다. 즉 목을 옆으로 움직이는 거북이와 활처럼 움직일 수 있는 거북이다. 분명히 이들 2 그룹은 목을 뒤로 움직이는 것은 따로 따로 발전시켰으며, 서로 다른 해결방식을 찾아냈다. 사실상, 쥬라기 후기에 발견되는 최초의 활모양의 목을 가진 거북이는 목을 뒤로 움직일 수 없었으며 그들의 후손이 그것을 가능하게 했다.
최초의 파충류에서부터 이궁류(diapsid)까지(see Evans, in Benton 1988, for more info)
- Hylonomus, Paleothyris (early Penn.)-- 위에 언급한 원시적인 amniotes 이다.
- Petrolacosaurus, Araeoscelis (late Pennsylvanian) -- 최초로 알려진 diapsids(이궁형). 2개의 섭유개구(temporal fenestra)가 존재한다. 턱근육은 크게 변하지 않았다. Hylonomus-style의 이빨을 가지고 있으며, 많고 작은 marginal teeth와 조금 큰 송곳니를 가지고 있다. 아직 고막은 없다.
- Apsisaurus (early Permian) -- 좀더 전형적인 이궁형 파충류이다. 송곳니가 없다. (Laurin, 1991)
GAP: no diapsid fossils from the mid-Permian.
- Claudiosaurus (late Permian) -- 초기의 diapsid 이며 여러 가지 이궁형 파충류의 새로운 특징을 가지고 있다. 그러나 아직도 원시적인 cervical vertebrae 와 unossified sternum을 가진다. 아마도 모든 아궁형 파충류(도마뱀, 뱀, 악아, 새)의 조상과 가까울 것이다.
- Planocephalosaurus(early Triassic) -- 좀더 도마뱀과 뱀과 비슷하게 변했다. 일부 두개골의 뼈와, 이빨, 발까락 뼈가 없어졌다.
- Protorosaurus, Prolacerta (early Triassic) -- 아마도 최초의 archosaurs에 속할 것이며, 이 계보가 공룡고 악어, 새들을 탄생시켰다. archosaurs의 사촌정도 될 것이다.
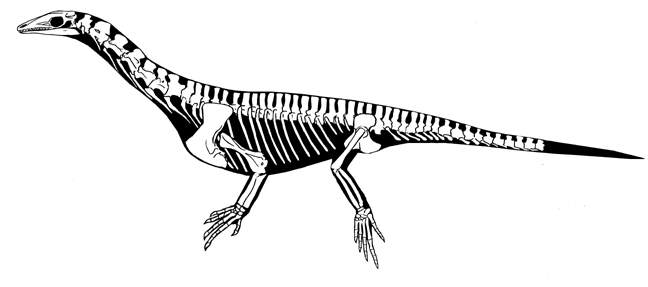 Protorosaurus
Protorosaurus
가장 오래된 archosauromorph, Protosaurus, 이며 20세기가 되기 전부터 알려졌다. 화석이 여러개 알려졌다. 길고 무거운 목이 Protorosaurus이 물개 같은 비율을 가지게 했다. 견갑골과 오훼골이 융합되어 있다. 꼬리의 hemal arches이 작아졌다.
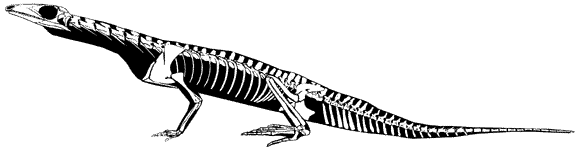
Prolacerta broomi
1935년도에 알려졌다. Prolacerta 은 최소한 5개의 표본이 알려졌고 이것은 부서지지 않고 완전한 것들이다. 원래 도마뱀과 비슷하다고 생각이 되었으며 이 원시적인 prolacertiform 은 초기의 archosauriforme, Proterosuchus와 매우 비슷하다. 목이 길고 꼬리의 base가 깊다.
- Proterosuchus (early Triassic) -- 최초의 archosaur.
- Hyperodapedon, Trilophosaurus (late Triassic) -- 초기의 archosaurs.
Some species-to-species transitions:
- De Ricqles (in Chaline, 1983) 은 초기 페름기의 파충류의 몇가지 과인 Captorhinus, Protocaptorhinus, Eocaptorhinus,와 Romeria의 점진적인 진화(다른 몇 개의 계보는 갑자기 나타난 것같이 보이기도 한다.)의 가능성을 말했다.)
- Horner et al. (1992) 은 최근에 매우 뛰어난 공룡의 전이화석을 백악기의 해안가였던 몬타나에서 매우 많이 발견했다. 아래의 것을 포함한다.:
- Styracosaurus 와 Pachyrhinosaurus 사이의 여러종류의 ceratopsids
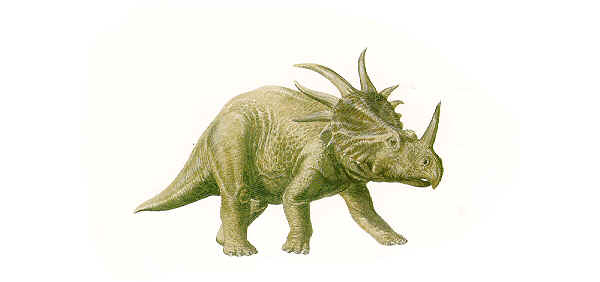 Styracosaurus
Styracosaurus- Lambeosaurus 와 Hypacrosaurus 사이의 lambeosaurids의 많은(50! 개) 전이화석들
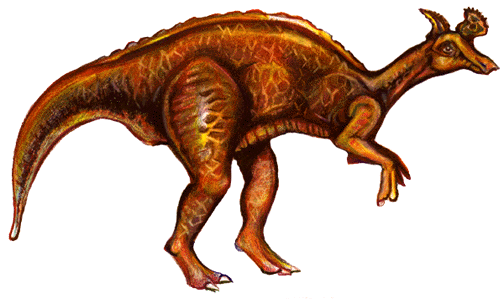 Lambeosaurus
Lambeosaurus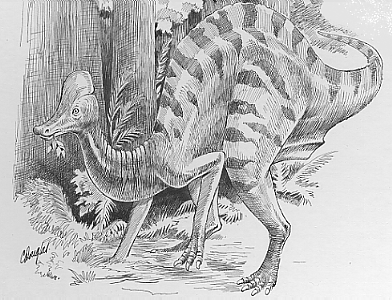 Hypacrosaurus
Hypacrosaurus- between Stegoceras와 Pachycephalosaurus 사이의 pachycephalosaurid 의 전이화석
 Stegoceras
Stegoceras
- Tyrannosaurus 와 Daspletosaurus사이의 tyrannosaurid 전이화석

 Daspletosaurus.
Daspletosaurus.이 모든 중간종들은 transitional animals 은 모두 같은 500,000년동안에 살았다. 이 사이트가 연구되기전에는 이러한 공룡 그룹은 Judith River Formation에서 알려진 것으로는 5백만년의 진화가 정지되고 갑자기 새로운 종이 나타나는 것으로 보였다. 50만년간에 해수면의 높이가 변화한 것으로 드러나서 잠시 Judith River Formation 이 물에 잠겼었고 공룡들의 숫자가 좀더 작은 지역 예를들어 몬타나같은 곳으로 몰리게 되었다. 다시 해수면이 낮아지자, 새로운 형태의 동물이 다시 Judith River로 퍼져나가게되었으며 이 때문에 중간화석이 몬타나지역에서만 발견되었다. 이것은 단속평형의 아주 뛰어난 예이며(예, 50만년은 매우 짧은 시기이며, 이정도면 단속되었다고 볼 수 있다) 이것은 또한 왜 중간 화석이 왜 작은 지역에서만 발견될 수 있고 다른 지역에서는 갑자기 나타나는가를 설명하는 좋은 예이다. (Horner et al., 1992)은 또한 Ianthosaurus, 의 발견으로 asynapsid 속 Ophiacodontidae 와 Edaphosauridae을 연결할 수 있다고 지적한다.(see Carroll, 1988, p. 367)
synapsid reptiles (단궁형 파충류)에서 포유류로
이것은 척추동물의 전이중에서 가장 잘 정리되어있다. 지금까지 이러한 계보는 속과 과 서열로 알려졌다. 종과 종간의 진하는 아직 알 수 없다. 그러나 속의 순서는 매우 완전하다. 각 그룹은 분명히 그 앞의 그룹과 그 뒤의 그룹과 명백하게 관련이 되어 있다. 그리고 그 순서가 매우 길기 때문에 마지막의 화석은 처음의 것과 비교하면 매우 변해 있다는 것을 알 게 된다. 최근에 Rowe(in Szalay et al., 1993)가 말했듯이 "시료를 취하는데 잘못을 저지르지 않고, 모든 가능한 특징에 대한 자료를 분석하면 [진화를 가정하지 않는 분류용 프로그램을 사용함], 매우 확실하고, 안정한 계통발생을 얻을 수 있으며 이것은 화석의 기록에 의한 분류와 일치한다. 비슷하게, Gingerich (1977)가 역시 말했는데 "현재 살아있는 포유류들이 서로 그룹이 잘 나누어지듯이, 화석기록은 파충류 조상에서부터 포유류까지 아주 자세하게 흔적을 보이고 있다. 좀더 자세히 알고 싶다면 Kermack's 뛰어나고 크지 않은 책 (1984)을 보거나, Kemp's의 좀더 자세하지만 약간 오래된 책 (1982), 과 Szalay et al.'의 최근 리뷰 논문을 모음(1993, vol. 1)을 보기바란다. .
이 리스트는 pelycosaurs (early synapsid reptiles)에서 시작하여 tehrapsid와 시노돈트에서 의심할 바 없는 포유류까지이다. 이러한 전이는 여러 가지 변화 즉 커지는 두뇌와 감각기관을 위한 두개골의 재편, 효과적으로 음식을 섭취하기 위해서 턱과 치아의 재모델링, 사지와 척추의 변화, 다리가 신체의 아래쪽으로 향하여 이동이 편하게 되는 것등이 포함된다. 아래의 것을 유심히 관찰해야 한다.
|
| ||
# | 초기 파충류 | 포유류 |
|
| ||
1 | 두개골에 섭유(fenestrae, 구멍)가 없다.l | 두개골(braincase)에 커다란 fenestra가 있다. |
2 | Braincase가 약하게 연결되어 있다. | Braincase attached firmly to skull |
3 | secondary palate가 없다. | 완전한 bony secondary palate(구개)가 있다. |
4 | 이빨이 분화되지 않았다. | 앞니(Incisors), 송곳니(canines), 앞어금니 (premolars), 어금니(molars) |
5 | Cheek teeth가 크라운이 없는 점모양이다. | Cheek teeth (PM & M) 크라운과 커습이 있다. (*) |
6 | 이빨이 연속적으로 교체된다. | 이빨이 기껏해야 한번 고체된다. |
7 | 이빨의 뿌리가 한개씩이다. | 어금니의 뿌리는 2개이다. |
8 | Jaw joint quadrate-articular | Jaw joint dentary-squamosal (*) |
9 | 아래턱(Lower jaw)은 여러개(3개)의 골판으로 이루어졌다. | 아래턱은 dentary bone 하나 뿐이다. |
10 | 귀의 뼈가 하나이다.(등골) | 귀의 뼈가 3가지이다. (등골, 침골, 추골) |
11 | 외부 콧구멍이 하나로 합쳐졌다. | 콧구멍이 분리되어있다. |
12 | 후두돌기가 한 개다. | 후두돌기가 2개이다. |
13 | Long cervical ribs | Cervical ribs tiny, fused to vertebrae |
14 | 허리부분에 갈비뼈가 있다. | 허리부분에는 갈비뼈가 없다. |
15 | 횡경막이 없다. | 횡경막이 있다. |
16 | 사지가 몸에서 옆으로 뻗어나왔다. (기어다닐 때 배가 땅에 닿는다) | 사지가 몸에서 아래쪽으로 뻗어나왔다. (기어다닐 때 배가 땅에서 떨어진다) |
17 | 견갑골이 단순하다. | 견갑골이 근육 때문에 크다 |
18 | 골반 뼈들이 서로 infused되지 않았다. | 골반뼈가 하나로 되어있다. |
19 | Two sacral (hip) vertebrae | Three or more sacral vertebrae |
20 | 발까락 뼈 숫자2-3-4-5-4 | 발까락 뼈 숫자 2-3-3-3-3 |
21 | 변온동물 | 항온동물 |
|
| ||
(*) 크라운이라는 것은 어금니의 경우밖에 노출되어 보이는 부분으로 치관(齒冠)이라고 부른다. 커습(cusp)은 이빨 상부의 갈 리는 표면 위에 날카롭게 솟아나온 부분을 말한다. 유태반류의 경우에는 송곳니 바로 뒤의 4개의 어름니를 앞어금니(pre-molar)라고 부르고 뒤의 3개의 어금니를 어금니라고 부른다. 앞어금니는 비교적 단순한 데 반하여 어금니는 먹이의 습성에 따라 변형되어, 그들의 먹이 습성을 잘 알려 준다.
(*) The presence of a dentary-squamosal jaw joint has been arbitrarily selected as the defining trait of a mammal.
- Paleothyris (early Pennsylvanian) -- 초기 captorhinomorph 파충류, 섭유개공(temporal fenestrae)이 아직 없다.
- Protoclepsydrops haplous (early Pennsylvanian) -- 최초의 단궁형(synapsid) 파충류. 섭유개구가 작게 하나 있으며, 주변의 뼈는 모두 intact하다. 단편으로만 남아있다. 양서류형태의 척추를 가지면 척추에는 신경돌기가 있다. (양서류에서 막 분리된 파충류임)
- Clepsydrops (early Pennsylvanian) --두 번째로 오래된 단궁형 파충류. 이 오래되고 매우 원시적인 pelycosaurs 그룹의 단궁류는 "ophiacodonts"라고 불린다.
- Archaeothyris (early-mid Pennsylvanian) -- 약간 후기의 ophiacodont. 작은 섭유개구가 있고 몇 개의 뼈(supratemporal) 크기가 작아졌다. Braincase는 여전히 두개골에 느슨하게 연결되어 있다. 이빨이 조금 분화하기 시작했다. 아직 지극히 원시적인 양서류/captorhinid의 턱, 두개골, 사지 자세의 특징을 유지하고 있다. 전형적인 파충류이지만 장골이 약간 크다.
- Varanops (early Permian) --섭유개구가 좀더 커졌다. Braincase의 바닥부분이 최초로 포유류비슷하게 되었다.으며 두개골의 나머지 부분과 좀더 강하게 붙어있다. (occiput more strongly attached). 아래턱은 턱의 근육조직이 처음으로 변하기 시작했다. (slight coronoid eminence). 신체는 좁고 깊어졌다: vertebral column more strongly constructed. Ilium further enlarged, lower-limb musculature starts to change (prominent fourth trochanter on femur). 이 동물은 좀더 움직이기 좋아하고 활동적이다. 조상이 되기엔 너무 늦게 나타나며, 아마도 사촌일 것이다.
- Haptodus (late Pennsylvanian) -- 최초로 알려진 sphenacodonts의 하나로, sphenacodont 특징을 보이는 반면 아직도 많은 ophiacodont의 특징을 가지고 있다. 후두골(직Occiput)이 아직 braincase에 붙어있다. 이빨은 이제 크기가 차이가 나서 송곳니가 커지고 전체적으로 적어지기 시작했다. 턱근육이 더 강해졌다. 척수부분과 연결부분이 포유류를 닮았다. 척수의 신경궁이 더 길어졌다. 힙이 2개가 아니라 3개의sacral vertebrae이 융합되어서 매우 강해졌다. 사지는 매우 잘 발달했다.
- Dimetrodon, Sphenacodon 혹은 유사한 sphenacodont (late Pennsylvanian to early Permian, 270 Ma) -- 좀더 발달된 pelycosaurs, 분명히 최초의 therapsids(다음)와 관련이 있다.

 Dimetrodon 은 분명히 사촌이지 조상은 아니다. 그러나 이것이 매우 완벽한 화석으로 알려졌기 때문에 sphenacodont anatomy에 좋은 모델이 된다. 중간크기의 섭유개공을 가진다. 이빨은 좀더 분화했으며, 작은 앞니, 2개의 크고 깊은 뿌리를 가진 윗 송곳니가 양쪽에 있고 작은 어금니(전어금니 어금니 모두 포함)로 이어진다. 이 이빨은 연속적으로 교체가 가능하다. 완전히 파충류의 턱 연결형태이다. 아래턱뼈는 여러개의 턱뼈로 이루어졌으며 최초의 고막과 관련된 증후를 보인다. 그러므로 이동물은 단지 땅바닥의 진동밖에는 느끼지 못했다. (이들은 파충류의 중이를 가지고 있다.). 척추는 아직도 긴 신경궁을 가지고 있다.(특히 Dimetrodon,등에 난 돛모양의 구조를 가짐), 그리고 튼튼한 운동 근육을 위해서 길고 전체를 가로지르는 등뼈를 가진다.
Dimetrodon 은 분명히 사촌이지 조상은 아니다. 그러나 이것이 매우 완벽한 화석으로 알려졌기 때문에 sphenacodont anatomy에 좋은 모델이 된다. 중간크기의 섭유개공을 가진다. 이빨은 좀더 분화했으며, 작은 앞니, 2개의 크고 깊은 뿌리를 가진 윗 송곳니가 양쪽에 있고 작은 어금니(전어금니 어금니 모두 포함)로 이어진다. 이 이빨은 연속적으로 교체가 가능하다. 완전히 파충류의 턱 연결형태이다. 아래턱뼈는 여러개의 턱뼈로 이루어졌으며 최초의 고막과 관련된 증후를 보인다. 그러므로 이동물은 단지 땅바닥의 진동밖에는 느끼지 못했다. (이들은 파충류의 중이를 가지고 있다.). 척추는 아직도 긴 신경궁을 가지고 있다.(특히 Dimetrodon,등에 난 돛모양의 구조를 가짐), 그리고 튼튼한 운동 근육을 위해서 길고 전체를 가로지르는 등뼈를 가진다.
 Sphenacodon
Sphenacodon
- Biarmosuchia (late Permian) -- A therocephalian -- 최초의 가장 원시적인 therapsid(수궁목, 獸弓目- 발달한 단궁류이다, 외비공이 등쪽에 있고 주둥이 끝에 가깝다. 페름기부터 쥬라기 초까지 있었다)이다. 여러 가지 원시적인 sphenacodontid 특징을 유지하고 있다 : 턱근육이 두개골 안으로 들어갔으며, 후두골이 평평해지고, palatal teeth가 있다. 새로은 특징은 다음과 같다 : 섭유개구가 좀더 커지고, 볼의 거의 전체를 차지한다 supratemporal bone(상섭유골) 은 완전히 없어졌다. 후두골의 판은 pelycosaurs가 약간 앞쪽으로 기울어진 것과는 달리 약간 뒤쪽으로 기울어져있고 아직 braincase에 강하게 결합되어 있다. 위턱뼈 (Upper jaw bone,maxillary) 은 확장이 되어서 코뼈에서 누골(lacrimal)을 분리시켜서 초기 파충류와 포유류의 중간이 된다. 그러나 구개의 서골(vomer)는 뒤쪽으로 발달되어 구개골의 아래쪽으로 확장되었다. 이것이 2차 구개의 최초의 단계이며, 시노돈트에서 보이는 것과 완전히 같은 것이다. 송곳니는 커지고치열에서 매우 중요한 역할을 하게 된다. 이빨이 다양하게 바뀌었다: 어떤 therocephalians (즉Scylacosaurus) 은 포유류처럼 한 개의 송곳니만 가지게 되며 커진다음에는 송곳니를 교체하지 않는다. Jaw hinge 는 모양과 형태에서 좀더 포유류를 닮았다. 턱의 근육조직은 좀더 강하다 (특히 포유류의 턱 근육). 양서류닮은 hinged upper jaw 은 결국 움직일 수 없게 되었다. 척추는 아직도 sphenacodontid를 닮았다. 이동방법에 있어서 좀더 근본적인 변화가있었는데, 좀더 자유롭게 움직이는 앞발, 좀더 똑바로 세워진 뒷발과, 포유류 닮은 대퇴부와 골반을 가지게 되었다. 원시적인 sphenacodontid 상박골을 가진다. 발까락은 포유류처럼 서로 같은 크기로 되었다. 목과 꼬리의 척추는 몸체의 척추와는 확연하게 다르게 되었다. 아마도 아래턱에 고막을 가지게되었을 것이다.
- Procynosuchus (latest Permian) -- 최초의 cynodont --매우 유명한 포유류를 닮은 수궁류이다.
 어떤 경우에는 이것이 최초의 포유류라고 생각되기도 한다.
어떤 경우에는 이것이 최초의 포유류라고 생각되기도 한다. - 아마도therocephalians에서 유래했을 것이다. 이것은 뚜렷한 2차 구개와 기타 여러 가지 두개골의 특징에서 판단할 수 있다. 매우 강한 턱 근육 때문에 섭유개공이 매우 크다. 커다란 구멍은 이제는 얇은 광대뼈에 의해서 둘러싸여져 있다. 2차 구개는 이제 vomer와 maxilla가 아닌 구개뼈로 구성이 되었으나(포유류처럼) 아직 작은 구개판이다. (이것은 자라면서 soft tissue와 더불어 완성된다). 아래쪽의 앞니는 6개에서 4개로 줄어들었다. (포유류는 3다.) 이빨은 아래턱의 전체의 3/4만 이빨이 나았고 나머지 부분은 작고 복잡한 연결부분(jaw hinge)을 형성하며 이부분은 아직도 파충류와 비슷하다. 척주는 드디어 포유류처럼 보이기 시작했으며, 최초의 2개의 vertebrae 는 머리의 움직임 때문에 약간 변형되었다. 그리고 허리부분의 척추는 갈비뼈가 없어지기 시작했으며, 가슴과허리부분이 분리되기 기능적으로 분리되기 시작했다. 견갑골(Scapula)은 모양이 변하기 시작했다. 힙에서 장골은 좀더 커지고 치골은 작아졌다. 아마도 횡경막이 있었을 것이다.
- Dvinia [also "Permocynodon"] (latest Permian) -- 또 하나의 초기의 cynodont이다. 이빨이 단지 찌르는 것이 아니라 새로운 모습으로 변하기 시작했다. 어금니에서 커습이 나타나기 시작했다. 섭유개공이 더욱 커졌다. braincase도 커지는 두뇌 때문에 변화가 있었다. 치골(dentary bone)이 이제 아래턱의 주요 뼈가 되었다. 초기 파충류에서 보인 아래턱의 다른 뼈들은 이제 jaw hinge에서 작은 뼈로 변하여 복잡하게 얽혀있다. 한 개의 후두돌기가 표면에서 2개로 갈라진졌다. Dvinia의 두개골의 뒷부분이 아직 안알려졌기 때문에 다음 단계에서 보이는 것들이 여기서 진화했는지는 알 수 없다. 신체대사의 속도는 약간 증가했으며 거의 항온 동물이었을 것이다.
- Thrinaxodon(early Triassic) --
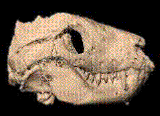 매우 발달한 "galesaurid" cynodont. 앞서 보인 여러 가지 cynodont으 특징이 더 발달했다. 섭유개공이 더 커지고, 턱근육이 더 커지고, 2개 구개가거의 완성되었고, 치아가 기능적으로 분리되었다. 즉 앞니(위에는 4개 아래는 3개), 송곳니, 그리고 씹기 위해서 커습을 가진 7-9개의 어금니가 있었다. 어금니가 서로 전어금니와 어금니로 구분되지는 않았지만 완전히 공간을 채운 것은 아니며, 한 개의 뿌리만을 가지고 있고 살아가면서 교체가 된다. Dentary 은 아직 크며, 작은 방골(方骨quadrate) 와 관절골 (articular)가 느슨하게 결합되어 있다. 등골(stpaes)은 이제 quadrate의 안쪽에 닿아있다. 포유류의 jaw hinge 의 모습이 처음으로 보이는데, 인대로 아래턱과 두개골의 squamosal을 연결하고 있다. 후두돌기는 이제 표면에서2개로 나뉘어져 있으니 포유류처럼 완전하게 분리되지는 않았다. 척추의 연결은 좀더 포유류와 비슷하고 허리의 갈비는 적어졌다. 견갑골은 새로운 포유류의 근육을 발달시켰다. 장골은 더 커졌고 4지는 똑바로 위를 향하게 되었고 파충류처럼 옆으로 퍼지지 않았다. 꼬리는 네발로 걷기에 편하도록 짧아졌으며 이동이 매우 빨라졌다. 발까락 뼈는 2.3.4.4.3,이며 이것은 파충류는 (2.3.4.5.4)이고 포유류는 (2.3.3.3.3)이므로 중간형태이고 포유류에 비해서 남는 뼈는 매우 작아졌다. 이 동물의 신체는 약간 말려 올려갔는데 아마도 열을 보존하기위해서였을 것으로 보이므로 안에서 몸을 뜨겁게 할 수 있었을 지도 모른다. 성숙한 것과 비성숙한 것이 모두 발견되었는데 이것은 부모가 새끼를 돌봐주었을 가능성을 보여준다. 허리부분의 갈비뼈가 줄어들었다는 것은 아마도 횡경막이 존재했을 것이라고 생각되며 항온을 위해서 산소를 들여마셨을 것이다. 고막이 드디어 생길 수 있는 가능한 곳 즉 아래턱, jaw hinge근처에 생겼으며 anglar bone의 넓은 판이지지해주고 있다. 이 동물이 드디어 소리를 들을 수 있다.이 소리는 고막을 eardrum을 통해서 아래턱의 작은 뼈 (관절골articular와 방골quadrate)로 전해지며, 이것은 다시 두개골의 stapes로 모아지고 이것은 다시 달팽이관으로 전달된다. 이것은 완전한 것은 아니며 특히 낮은 주파수의 소리만을 민감하게 들을 수 있지만 전혀 없는 것보다는 훨씬 낫다. Cynodonts는 느슨한 관절골과 방골을 개발하여 전달되는 소리에 의해서 진동할 수 있고 턱뼈의 연결부분으로도 작용할 수 있게 되어있다. 이것은 곧 포유류의 턱뼈 연결로 바뀌게 된다. 초기쥬라기의 초기의 포유류는 낮은 주파수를 들을 수 있는 귀와 이중의 턱연결부분을 갖는다. 중기 쥬라기에서는 포유류는 파충류식의 연결방식을 버리게 되고(배에서는 나타난다) 2개의 뼈는 중이로 이동하게 되고 매우 작고 높은 주파수의 소리에도 민감해진다.
매우 발달한 "galesaurid" cynodont. 앞서 보인 여러 가지 cynodont으 특징이 더 발달했다. 섭유개공이 더 커지고, 턱근육이 더 커지고, 2개 구개가거의 완성되었고, 치아가 기능적으로 분리되었다. 즉 앞니(위에는 4개 아래는 3개), 송곳니, 그리고 씹기 위해서 커습을 가진 7-9개의 어금니가 있었다. 어금니가 서로 전어금니와 어금니로 구분되지는 않았지만 완전히 공간을 채운 것은 아니며, 한 개의 뿌리만을 가지고 있고 살아가면서 교체가 된다. Dentary 은 아직 크며, 작은 방골(方骨quadrate) 와 관절골 (articular)가 느슨하게 결합되어 있다. 등골(stpaes)은 이제 quadrate의 안쪽에 닿아있다. 포유류의 jaw hinge 의 모습이 처음으로 보이는데, 인대로 아래턱과 두개골의 squamosal을 연결하고 있다. 후두돌기는 이제 표면에서2개로 나뉘어져 있으니 포유류처럼 완전하게 분리되지는 않았다. 척추의 연결은 좀더 포유류와 비슷하고 허리의 갈비는 적어졌다. 견갑골은 새로운 포유류의 근육을 발달시켰다. 장골은 더 커졌고 4지는 똑바로 위를 향하게 되었고 파충류처럼 옆으로 퍼지지 않았다. 꼬리는 네발로 걷기에 편하도록 짧아졌으며 이동이 매우 빨라졌다. 발까락 뼈는 2.3.4.4.3,이며 이것은 파충류는 (2.3.4.5.4)이고 포유류는 (2.3.3.3.3)이므로 중간형태이고 포유류에 비해서 남는 뼈는 매우 작아졌다. 이 동물의 신체는 약간 말려 올려갔는데 아마도 열을 보존하기위해서였을 것으로 보이므로 안에서 몸을 뜨겁게 할 수 있었을 지도 모른다. 성숙한 것과 비성숙한 것이 모두 발견되었는데 이것은 부모가 새끼를 돌봐주었을 가능성을 보여준다. 허리부분의 갈비뼈가 줄어들었다는 것은 아마도 횡경막이 존재했을 것이라고 생각되며 항온을 위해서 산소를 들여마셨을 것이다. 고막이 드디어 생길 수 있는 가능한 곳 즉 아래턱, jaw hinge근처에 생겼으며 anglar bone의 넓은 판이지지해주고 있다. 이 동물이 드디어 소리를 들을 수 있다.이 소리는 고막을 eardrum을 통해서 아래턱의 작은 뼈 (관절골articular와 방골quadrate)로 전해지며, 이것은 다시 두개골의 stapes로 모아지고 이것은 다시 달팽이관으로 전달된다. 이것은 완전한 것은 아니며 특히 낮은 주파수의 소리만을 민감하게 들을 수 있지만 전혀 없는 것보다는 훨씬 낫다. Cynodonts는 느슨한 관절골과 방골을 개발하여 전달되는 소리에 의해서 진동할 수 있고 턱뼈의 연결부분으로도 작용할 수 있게 되어있다. 이것은 곧 포유류의 턱뼈 연결로 바뀌게 된다. 초기쥬라기의 초기의 포유류는 낮은 주파수를 들을 수 있는 귀와 이중의 턱연결부분을 갖는다. 중기 쥬라기에서는 포유류는 파충류식의 연결방식을 버리게 되고(배에서는 나타난다) 2개의 뼈는 중이로 이동하게 되고 매우 작고 높은 주파수의 소리에도 민감해진다. - Cynognathus (early Triassic, 240 Ma; suspected to have existed even earlier) -- (단궁아강 수궁목의 대표적인 동물) 드디어 발달된 시노돈트이다. 섭유개공이 좀더 커졌고 이빨이 좀더 분화가 되었고 어금니가 커습이 제대로 되어서 드디어 아구가 맞아서 맞아 음식을 짜르는데 잘 사용될 수 있다. 이빨을 교체하는 속도가 줄어들었으며 뿌리는 하나이지만 모양은 포유류의 것과 비슷하게 바뀌었다. Dentary 가 여전히 커지고 아래턱의 90%의 근육이 있는 부분을 차지하게 되었다. JAW JOINTS가 두 개 생겼다. 포유류의 방식과 파충류의 방식인데, 새로운 뼈의 연결은squamosal (두개골에 있는)과 surangular bone (아래턱에 있는)사이에서 일어나고 다른 턱의 연결방식은 매우 작아져서 중이에 가까운 곳에 놓이게 되었다. 갈비뼈는 좀더 포유류처럼 되었다. 견갑골은 좀더 포유류처럼 되었으며, 사지는 몸의 아래쪽에 있게 되었다. 발자국에서 털의 증거가 나타날 수 있다.
- Diademodon (early Triassic, 240 Ma; same strata as Cynognathus) -- 섭유개공은 턱의 근육 때문에
 더욱 커졌으며, 진정한 뼈로 이루어진 2차 구개가 포유류와 같은 방식으로 생겨났으나 이것이 뒤로 완전히 확장되지는 않았다. 갑개골이 코안에 있었을 것으로 보인다. 치아도 계속 변했는데 치아를 바꾸는 속도가 매우 늦어졌고, 어금니는 좀더 커습이 알맞게 되었고 서로 잘 들어 맞았다. 아래턱은 완전히 치골로 거의 이루어지게 되었으며 다만 작은 힌지에 관절이 있을 뿐이다. 아직도 이중으로 턱이 연결되며, 갈비는 여전히 허리부분에서 짧아지고 이것으로 횡경막의 기능과 이동은 더욱 발달했을 것이다. 발까락의 뼈가 드디어 포유류의 것과 같아졌다.(2.3.3.3.3)
더욱 커졌으며, 진정한 뼈로 이루어진 2차 구개가 포유류와 같은 방식으로 생겨났으나 이것이 뒤로 완전히 확장되지는 않았다. 갑개골이 코안에 있었을 것으로 보인다. 치아도 계속 변했는데 치아를 바꾸는 속도가 매우 늦어졌고, 어금니는 좀더 커습이 알맞게 되었고 서로 잘 들어 맞았다. 아래턱은 완전히 치골로 거의 이루어지게 되었으며 다만 작은 힌지에 관절이 있을 뿐이다. 아직도 이중으로 턱이 연결되며, 갈비는 여전히 허리부분에서 짧아지고 이것으로 횡경막의 기능과 이동은 더욱 발달했을 것이다. 발까락의 뼈가 드디어 포유류의 것과 같아졌다.(2.3.3.3.3)
- Probelesodon (mid-Triassic; South America) -- 섭유개공이 더욱 커졌다. 2차 구개는 좀더 길어졌지만 아직 완성되지는 않았다. 어금니는 포유류처럼 2개의 뿌리를 갖게 되었다. 콧구멍이 2개가 분리되었으며, 나중에 생긴 턱 연결이 강해졌다. 허리부분의 갈비가 완전히 없어졌으며, 흉부의 갈비뼈도 포유류처럼 되었다. 척추의 연결이 포유류같이 되었고 힙과 대퇴부도 포유류처럼 변했다.
- Probainognathus (mid-Triassic, 239-235 Ma, Argentina) --두뇌가 매우 커졌고 두개골이 다양한 크기로 되었다 : 두정공(pineal foramen,"third eye")이 닫혀있다, 몇 개의 두개골 판이 합쳐졌다. 광대뼈가 얇아지고, 눈구멍의 아래가가라 앉았다. 이빨에는 추가적인 커습이 있다. 아직도 이중의 턱연결방식을 사용하고 있으며, 아직도 허리부분의 갈비뼈가 있으나 매우 작다. 파충류의 흉부의 늑골판은 거의 없어졌다. 포유류의 발까락 뼈의 수와 같이 되었다.
- Exaeretodon (mid-late Triassic, 239Ma, South America) -- (Formerly lumped with the herbivorous gomphodont cynodonts.) 푸유류의 턱 형태이며, 고막을 지탱하여 준다. 앞니가 3개뿐이며 흉부의 늑골판은 완전히 없어졌다. 좀더 포유류 닮은 힙을하고 있으며 이동과 호흡의 조화를 갖추기 시작한 최초의 단계로 보인다. 이것은 사촌이지 조상화석은 아닌 것으로 보이며, 새로운 포유류의 특징을 가지고는 있지만 포유류의 특징이 아닌 이빨을 하고 있다.
GAP 3천만년전의 트라이아스기 후기에 간격이 있으며 시기는 대강 2390만년 - 2080만년사이이다. 현재까지 초기의 포유류의 화석은 단 하나만 발견되었다. 다음에 포유류가 나타날 때는 이미 tritylodontids 와 trithelodontids 이 나타난 뒤이며 결국 이들간의 포유류와의 상대적인 위치가 논쟁거리이다. 최근 발견으로는 trithelodontids이 좀더 포유류를 닮았으며 아마도 tritylodontids가 곁가지가 아닌가 생각하고 있다 (see Hopson 1991, Rowe 1988, Wible 1991, and Shubin et al. 1991). 이들 모두가 벌써부터 거의 포유류에 매우 가깝고 단지 마지막 변화인 턱의 연결과 중이에 대한 변화만이 남았음을 기억하기 바란다.
- Oligokyphus, Kayentatherium (early Jurassic, 208 Ma) -- 이들은 삼괴치목( tritylodontids-앞어금니와 어금니의 구분이 뚜렷하지 않으며 전형적인 것은 송곳돌기가 3줄로 배열한다. )에 속하며 발달한 cynodont이다. 얼굴형태가 포유류와 더욱 비슷해 졌다. 눈 구멍 부분과 광대뼈 부분이 포유류 비슷해졌다. 완전한 뼈로 이루어진 2차 구개가 있다. 어금니를 교체하지만 포유류의 형태의 완전히 이가 맞는 것은 아님 (일부 시노돈트가 앞서 가지고 있던 특징임) 신체의 뼈구조는 단궁류와 너무 닮아있다. 이중의 턱 joint를 가지고 있으며, 좀더 유연한 목을 가지고 있고 후두돌기를 가지고 있다. 꼬리 척추부분은 포유류처럼 단순해졌고, 견갑골은 포유류처럼 되었다. 앞발은 신체의 바로 아래에 있게 되었다. 골반과 뒷발 근육에서 많은 변화가 있었는데 이 동물의 근육조직과 운동은 완전히 포유류의 것과 동일하다. 아마도사촌 화석으로 생각되며, Oligokyphus 이 좀더Kayentatherium보다 원시적이다. 비록 삼괴치목이 쥬라기 말기에 다양하게 퍼지긴 했지만 이들이 포유류의 조상인지 혹은 포유류와는 관련이 없는 지나치게 특수화된 offshoot 그룹인지 아닌지는 아직 일치되지는 않았다.
- Pachygenelus, Diarthrognathus (earliest Jurassic, 209 Ma) -- 이들은 삼괴치목에 속하며 약간 다르게 발달한 시노돈트 그룹이다.새로 발견된 내용으로 이들이 매우 포유류와 가깝다는 것이 밝혀졌다. 비강이 커졌으며, 유스타키오관이 귀와 인두를 연결하고 눈뒤의 bar는 없어졌다. 뿌리가 하나뿐인이빨들이 순차적으로 새로 나게 된다. 이 그룹은 2개의 뿌리를 가진 이빨을 발전시키기 시작했다. Pachygenelus 에서 한 개의 뿌리를 가진 어금니가 기저에서 2개로 갈라지기 시작했다. Pachygenelus 는 또한 포유류같은 이빨의 에나멜층이 있으며 이빨아 아래위가 잘 들어맞는다. 턱 연결부분이 이중으로 되어있고 두 번째의 연결이 치골-squamosal (surangular대신)으로 되어있어 완전히 포유류의 것과 동일하다.파충류의 턱연결방식이 존재하지만 거의 듣기만을 위해서 기능한다. 후치골(postdentary bones)은 중이근처에서 좀더 작은 막대형으로 되어서 고주파수의 음도 들을 수 있게 되었다. 좀더 포유류 닮은 척추와 자유롭게 움직이는 목을 가지고 있다. 힙은 좀더 포유류를 닮았고, 장골과 대퇴부가 거의 포유류의 것과 유사하게 되었다. 상당히 잘 움직이는 포유류 스타일의 어깨를 가지고 있다. 아마도 이동과 호흡이 서로 조화를 이룰 것이다. 이들이 조상화석은 아니지만 매우 가까운 사촌으로 보인다. (진정한 조상화석은 아마도 트라이아스기 말기에 발생했을 것으로 보인다.)
- Adelobasileus cromptoni (late Triassic; 225 Ma, west Texas) -- 최근에 발견된 화석이며 중기 혹은 말기의 트라이아스갭에서 발견된 원포유류(proto-mammal) 화석이다. 현재로선 가장 오래된 포유류이며, 아직 두개골만이 발견되었다. Adelobasileus두개골의 특징은 달팽이관을 싸는 것이 시작되었다는 것인데 이것은 포유류가 아닌 시노돈트에서 Liassic mammals로 전이되는 것을 보여준다. (Lucas & Luo, 1993). 이 화석은 미국의 서부의 포유류의 화석을 연구하지 않던 지층에서 나왔다. 그리고 이들 화석이 알져진삼괴치목과 trithelodonts보다 전에 생긴 것으로 연대측정이 되었다. Adelobasileus 은 삼괴치목이나 trithelodont에서 갈라져 나왔거나 아니면 이들이 서로 포유류의 조상이라고 생각된다.
- Sinoconodon (early Jurassic, 208 Ma) -- 다음으로 알려진 오래된 원포유류. 눈구멍은 이제 완전히 포유류의 모양 (closed medial wall)이다. 후뇌가 발달했다. 포유류처럼 영구 어금니가 생겼으나 다른 이빨은 여러번 갈 수 있게 되었다. 포유류 방식의 턱의 연결은 매우 강하고 치골돌기는 squamosal의 구멍에 아주 잘 맞는다. 이 정교한 턱의 연결은 이제 완전히 포유류로 되게 했지만 아직도 작지만 파충류의 턱 연결을 가지고 있다.
- Kuehneotherium (early Jurassic, about 205 Ma) -- 약간 나중에 생긴 원포유류로 종종 이들을 최초의 pantothere (원시적인 태반-타입의 포유류)라고 부른다. 이빨과 두개골은 태반포유류와 매우 비슷하다.3개의 주요 커습이 좀더 발달된 태반 포유류나 유대류에서 같이 서로 막아주면서 삼각형을 이루면서 잘 갈아준다. 그럼에도 불구하고 아직도 2중의 턱 연결구조를 하고 있다.
- Eozostrodon, Morganucodon, Haldanodon (early Jurassic, ~205 Ma) -- 초기 원 포유류의 그룹이며 "morganucodonts"라고 불린다. 2차 구개의 재편과 braincase의 바닥의 변화가 계속 일어났다. 이제 거의 포유류를 닮았다. 진정한 포유류의 이빨을 하고 있다. 어금니가 드디어 단순한 전 어금니와 복잡한 기능을 하는 어금니로 분화가 일어났다. 그리고 이빨을 단 한번 교체할 수 있을 뿐이다. 삼각의 커습을 가진 어금니를 가지고, 앞니가 줄어드는 방향에서 반대로 오히려 4개로 늘어났다. 매우 작은 파충류의 턱연결을 하고 있다. 처음에는 이것이 단공류의 시조로 생각이 되었으나 지금은 현대의 모든 포유류(단공류, 유대류, 유태반류)의 조상으로 생각이 되고 있다.
- Peramus (late Jurassic, about 155 Ma) -- "eupantothere" (좀더 발전된 태반타입의 포유류)이다. 알려진 것중 유태반류와 유대류의 가장 가까운 친척이다. 3돌기치목의 어금니와 좀더 세련된 커습을 가지고 있다. 이 화석은 오직 이빨만이 알려졌으나 이와 유사한 eupantotheres (e.g. Amphitherium)에 근거하여 생각해 볼 때 이제 파충류의 턱 연결은 완전히 없어졌고 포유류의 3개의 뼈로 이루어진 중이가 생겼으며 결과적으로 뛰어난 고주파 청음 능력을 가지게 되었다. 8개의 어금니를 가지고 있으며 이것은 다른 eupantotheres 보다 적으며 최초의 포유류의 7개에 근사하게 되었다. 또한 어금니에 커다란 talonid가 있으며, 어금니의 크기도 초기 유태반류처럼 크다. 최초로 갈 수 있는 능력이 생겼다.
- Endotherium (very latest Jurassic, 147 Ma) -- 발달된 eupantothere. 완전히 tribosphenic 어금니이며 잘 발달된 talonid를 가진다. 한 개의 개체밖에 발견되지 않았다. 아시아에서 발생한 것을 보인다. :최근의 아시아의 화석에서 tribospheric 어금니가 거기서 진화한 것으로 추정된다.
- Kielantherium 와 Aegialodon (early Cretaceous) -- 좀더 발달한 eupantotheres 이지만 치아만 알려졌다. Kielantherium 은 아시아에서 알려졌고, 유럽의 Aegialodon보다 오래된 지층에서 발견되었다. 이들은 모두 아래쪽 어금니에 talonid를 가지고 있다. 새로운 커습 (protocone)이 윗 어금니에서 진화되었다. 백악기 중기에, 새로운 tribosphenic molar동물이 북미대륙으로 이동했다. (북미 대륙이 아시아와 연결되어 있었다.)
- Steropodon galmani (early Cretaceous) -- 최초의 분명한 단공류(monotreme), 1985발견
- Vincelestes neuquenianus (early Cretaceous, 135 Ma) -- 아마도 유태반류의 포유류이며 약간의 유대류의 특징을 가지고 있다. 몇 개의 좋은 두개골 화석이 남아있다. 유태반류타입의 braincase와 돌돌말린 달팽이관을 가지고 있다. 두개골안에서 동맥과 정맥이 단공류와 유태반류의 혼성으로 흐르고 있으며 시노돈트의 두개골밖에서 흐르는 것과 상동으로 보인다. (Rougier et al., 1992)
- Pariadens kirklandi (late Cretaceous, about 95 Ma) -- 최초의 명백한 유대류. 오직 이빨만이 알려졌다.
- Kennalestes and Asioryctes (late Cretaceous, Mongolia) -- 조그맣고 가느다란 동물이다. 눈구멍이 뒤에 열려 있고 고막을지지하는 단순한 링이 있다. 원시적인 유태반류 타입의 두뇌를 하고 있으며 커다란 후각의 구(olfactory bulbs)를 가지고 있다. 기초적이고 원시적인 tribosphenic 이빨 패턴을 보인다. 송곳니는 이제 두 개의 뿌리를 가지고 있다. 아직 치골이외의 흔적 (즉 coronoid)가 있지만 이것을 제외하면 좋은 치골의턱을 가지고 있다. 아마 모든 유태반류의 조상을 일 것이라고 Carroll (1988)이 말했다.
- Cimolestes, Procerberus, Gypsonictops (very late Cretaceous) -- 원시적인 북미의 유태반류이며 아직 기본적인 이빨 패턴을 보여준다.
그러므로, 백악기 후기까지 3 그룹의 현대의 포유류가 같이 생겨났다. 단공류와 유대류와 유태반류이다. 유태반류는 동아시아에서 생겨나서 백악기 말기에는 미국으로 건너갔다. 백악기 말기에 공룡이 거의 사라지고 나서 유태반류와 유대류가 조금씩 다양화되기 시작했다. 효신세에서는 이런 다양성이 매우 가속화되었다. 예를들면 효신세 중기에는 유태반류의 화석은 영장류 닮은 동물 (Purgatorius - known only from a tooth, though, and may actually be an early ungulate), 좀더 잘 갈기 위해서 약간 평평해진 어금니를 가진 초식동물 턱(Protungulatum, probably an early ungulate)과 곤충을 잡아먹는 동물 (Paranyctoides)이 나타났다.
어떤 것이 최초의 포유류인지는 약간 주관적이다. 우리는 약간 inflexible 한 분류 체계로 이 점진적인 시리즈를 나누어 보았다. 진정한 파충류에서 포유류로의 중간 그룹이 생겼었으며 이들은 점차적으로 포유류의 특성을 획득하여 결국 우리가 포유류로부터 파충류까지 이어지는 선을 차지하게 되었다. 예를들어 Pachygenulus 와 Kayentatherium 는 모두 파충류보다는 포유류에 가깝다. 하지만 그들은 모두 파충류로 불린다.
이궁형 파충류에서 조류
1800년대 중반에서는 이것은 척추동물에서 가장 큰 간격이었다. 아무 중간화석이 알려지지 않았었기 때문이며, 이 두 그룹은 불가능할 정도로 완전히 달라보였기 때문이다. 그러다가 1861년 시조새(Archeopteryx)의 기가막힌 화석이 발견되었는데 이는분명히 두 그룹이 서로 연결될 수 있음을 보여주었다. 그때 이후로 다른 파충류와 조류의 연결고리가 발견되었다. 그러나 전체적으로 본다면아직도 많은 중간화석이 발견되지 않았다. 나는 Mononychus (이 새는 날아다니는 것보다는 땅을 파고 살았다. 현대의 새와 상당히 떨어져 있다.)는 포함시키지 않았다. Feduccia (1980) 와 Rayner (1989)를 살펴보면 어떻게 날기가 진화되었는지 다루고 있으며, Chris Nedin's 은 뛰어난 Archeopteryx FAQ를 제공했다.
- Coelophysis (late Triassic) -- 최초의 theropod 공룡의 하나. Theropods는 일반적으로 새와 비슷한 골격을 보인다. (긴 사지, 속이 빈 뼈, 발에 앞발까락이 3개이고 뒤에 발까락이 1개, 긴 장골(腸骨,ilium, 골반을 형성하는 뼈의 하나). Compsognathus 같은 쥬라기의 theropods는 부분적으로 새와 매우 비슷하다.
- (theropoda : 수각獸脚아목 : 삼첩기, 백악기, 육식을 하며, 앞다리는 극히 짧다. 따라서 보행애 사용하지 않았던 것으로 추정된다. 이들 중에는 지금까지 알려진 가장 큰 육상 육식 동물이 있었고, 티라노 사우르스, 알로사우르스, 메갈로사우르스, 시피노사우르스가 포함된다. 용반목의 수각아목임)
 이 동물은 Rioarribasauruses (처음에는 Coelophysis라고 불렸슴)라고 불린다. Rioarribasarus은 archosaurs에서 진화했으나 삼첩기의 끝이 되서야 나타난다.
이 동물은 Rioarribasauruses (처음에는 Coelophysis라고 불렸슴)라고 불린다. Rioarribasarus은 archosaurs에서 진화했으나 삼첩기의 끝이 되서야 나타난다.
- Deinonychus, Oviraptor, 그리고 다른 발전된 수각류 (쥬라기 말기, 백악기) -- 양발로 걸어다니는 점도 진보적인 육식성의 수각류, 좀더 커지고, 새와 비슷한 뼈의 구조를 하고 있다 : 반월 모양의 손목관절, 뼈로 이루어진 흉골 (가슴판, 새의 흉골은 앞가슴의 판을 말한다.) 긴팔, 반대방향의 치골. 분명히, 날지는 못해도 매우 잘 달렸다. 쇄골이 있기는 했지만 이것은 가끔은 새처럼 fused되어있다. Says Clark (1992): "새와 테로포드 공룡 예를들어 Deinonychus 사이에는유사성에 있어서 매우 놀랍고 여러개의 skeleton에서 많은 부분에서 유사하기 때문에 새의 친척들의 그룹이 테로포드 공룡에 속한다는 주장을 하지 않을 수 없다"라고 했으며, 여기에 소개한 특정 화석이 조상화석은 비록 아니지만 이들은 시조새의 뒤에 나타난다.
- Lisboasaurus estesi & other "troodontid dinosaur-birds" (mid-Jurassic) -- 새를 닮은 theropod 파충류이다. 새를 닮은 치아 (즉, 여기서 치아는 초기의 이빨을 가진 새를 말한다. 현대의 새는 이빨이 없다.). 이들이 실제로 조상이 될 것이다.
GAP: 어떤 것이 시조새의 조상인지 그리고 어떻게 날개깃이 발전되었는지 알려져 있지 않다. 최초의 새의 진화는 아마도 작은 숲을 오르내리는 동물이 다음에는 숲을 날아다니는 동물로 진화한 것으로 보인다. 이러한 상황이라면 화석이 거의 남지 않는다 (작은 동물 + 산성의 숲의 토양 = 잔해가 없음) Archeopteryx 자신은 우리가 얻을 수 있는 최상의 것이다. 여러개의 표본이 날개의 자국까지 선명하며, 이것은 파충류와 조류에 모두 관련이 있으며 이러한 진화가 가능함을 보여주고 있다.
- 시조새의 조상으로 가능한 것으로는 Protoavis (Triassic, ~225 Ma)가 있다. -- 초기의 새인지 아닌지 매우 논란이 많은 화석이다. 불행하게도 새와 관련이 있는지 충분하게 알 수 있을 정도의 화석이 발견되지 않았다.
- Archeopteryx lithographica (Late Jurassic, 150 Ma) -- 여러 가지 알려진 표본으로부터 파충류와 조류의 특징을 모두 모자이크 해서 가지고 있다는 것을 보여주고 있다. 즉 두개골과 skeleton (이빨, 척추, 흉골, 갈비뼈, 골반, 꼬리, 발까락, 발톱(claw)와 일반적으로 융합되지 않은 뼈들)은 파충류를 닮았으며, 새의 특징은 가슴의 창사골(날기위한 근육이 붙어있기 위한 것이며, 몇몇 공룡도 이것을 가지고 있음을 기억하자), 변형된 앞발과 잘 달리기 위한 발, 그리고 날기 위한 날개깃이 있다. Archeopteryx 는 나무에서 나무로 뛰어 이동할 수는 있었지만 지상에서 날지는 못했을 것이다. 왜냐하면 이들은 날기위한 근육이 붙어있을 만한 용골돌기(keel)이 있는 가슴뼈가 부족하고, 현대의 새와 비교해서 어깨가 약하다. 아마도 현대의 새의 직접적인 조상은 아닐 것으로 보인다. (Wellnhofer, 1993)
- Sinornis santensis ("Chinese bird", early Cretaceous, 138 Ma) -- 최근에 발견된 원시적인 새이다. 새의 특징으로는 : 짧은 몸통, 발까락의 발톱, 날기에 적합한 어깨, 강한 깃뼈, 강하게 말려있는 손목, 짧은 손 (이러한 특징은 이것이 시조새보다 훨씬 잘 날 수 있음을 보여준다.) 파충류의 특징으로는 이빨, 위(stomach)의 갈비뼈, 완전히 융합되지 않은 손의 뼈, 파충류 닮은 융합되지 않은 골반(이러한 특징이 날개를 방해하지는 않는다.) 중간적인 특징으로는 족근골이 어느정도 융합되었고, 중간 크기의 흉부의 용골돌기가 있으며 중간크기의 꼬리 (8개의 척추뼈가 있다.)가 끝에 pygostyle가 융합된 형태로 있다. (Sereno & Rao, 1992).
- "Las Hoyas bird" 또는 "Spanish bird" [아직 이름이 없음, 초기 백악기, 131 Ma) -- 최근에 발견된 작은 숲속의 비행자 "little forest flier". 아직 파충류 닮은 골반과 다리를 하고 새를닮은 어깨를 하고 있으며 꼬리는 중간 크기이고 끝이 융합되어 있다. 작은 좀털 (down)이 발견되었는데 이것으로 이 동물이 항온동물일 것으로 추측된다. (Sanz et al., 1992)
- Ambiortus dementjevi (early Cretaceous, 125 Ma) -- 세 번째로 알려진 "작은 숲속의 나는 새"는 1985년 발견되었으며, 매우 단편으로 갈라진 화석이다.
- Hesperornis(해거름새), Ichthyornis(물고기새목), and other 백악기의 다이빙하는 새들 -- 이 계통의 새는 현대의 가마우지 처럼 다이빙을 하도록 특수화되었다. 이들은 짠물이 있는 해안을 따라서 살았으며, 알려진 많은 화석이 있다. 신체의 뼈는 좀더 날기에 적합하게 진화했으며 (치골이 융합되었고, 손의 뼈가 융합되었고, 꼬리가 짧고 융합되었다.) 아직도 socketed teeth를 가지는데 이것은 파충류의 특성이다.
 hesperonis
hesperonis  Ichthyornis
Ichthyornis
[Note: 고전적인 병아리의 배야의 연구에 의하면 병아리의 부리는 이빨을 발달시킬 수 있을 것으로 보인다. 병아리의 유전자에는 이빨을 만드는 유전자가 아직 있다. 그리고 분자 유전학의 자료로볼 때 크로코다일 악어가 새와 가장 유사한 파충류이다. ]
 중간화석에 대한 정리 -- 1
중간화석에 대한 정리 -- 1