분자 서열 증거는 모든 생명체의 계통적인 친척관계에 대한 가장 인상적이고, 반박할 수 없는 증거를 제공합니다. 분자 서열의 본질상, 공통조상에 대한 예측이 실험적인 관찰과 실제로 얼마나 잘 맞는가를 보여줄 수 있는, 놀라운 확률 계산을 가능하게 합니다. 공통조상은 실험적으로 관찰된 분자 증거에 근거를 둔 가설에서 직접적으로 연역되는 것입니다. 게다가, 생물학의 분자적 메카니즘과 구조와 대진화 이론이 결합하여 매우 특이적이고, 색다르고, 검증할 수 있는 생물분자적 예측을 제공합니다.
계통발생적인 친척관계를 결정하는 분자서열 증거들은 여러 가지 분류로 나뉠 수 있으며 서로 각각의 독립적인 증거를 가지고 있습니다. 기능성 및 비기능성 유전자의 특성 모두가 일상적으로 비교됩니다. 기능성 인자의 연구에는 rRNA, 편재 유전자, 그리고 미토콘드리아 DNA의 비교가 포함되며, 비기능성 인자의 연구에는 가유전자, 내재 레트로바이러스 유전자, 및 모바일유전 인자(예를들어 인트론, 트랜스포존, 혹은 레트로인자)가 포함됩니다.
예측 4.1: 기능이 있는 분자에 의한 증거 - 단백질 중복성
분자 서열 연구에 의해 주어진 공통 조상을 지지하는 내용을 다음과 같은 연역적인 논증으로 바꾸어질 수 있습니다. 이 논증은, 유사성이 친척관계를 암시한다고 직접적인 결론을 내릴 수 있는 유일한 사례이기 때문에 이 FAQ에서 매우 독특한 것입니다. 이 결론은 특정한 상황에서 생물학적 구조의 유사성에 의존하고 있습니다. 즉, 서로 다른 종들의 편재 유전자간의 유사성이 관찰된다는 것입니다.
아래의 논의는 약간은 기술적입니다. 그러므로 우선 연역적인 주장의 개요를 제시하며, 이것을 통해서 논리적인 흐름을 쉽게 파악할 수 있을 것입니다. 우선 주장의 각 전제를 서술하고 그 다음에 결론이 나오며 그 뒤로 토의가 이어집니다.
주장의 요점:
(P1) 편재(遍在) 유전자(Ubiquitous genes) : 기본적인 생명 기능을 수행하기 때문에 모든 생명을 가진 유기체에 존재하는 유전자들이 있으며 이러한 유전자를 편재 유전자라고 한다. (역주 : 일반적으로 편재라고 하면 한쪽으로 치우쳐 있다는 의미로 偏在라는 한자를 사용합니다. 하지만 위의 용어는 골고루 분포한다는 의미로 한자가 다릅니다.)
(P2) 편재 유전자들은 종-특이적인 표현형과 관련이 없다. : 편재 유전자는 서로 다른 종들의 종만의 특별한 기능과는 전혀 관계가 없다. 예를 들어, 당신이 박테리아이건, 인간이건, 개구리이건, 고래이건, 벌새이건, 달팽이이건, 곰팡이이건, 말미잘이건 전혀 문제 없이 이러한 편재유전자를 가지고 있으며, 당신이 무엇이건 간에 동일한 기본적인 생물학적 기능을 수행한다.
(P3) 편재 유전자의 분자 서열은 기능적으로 중복(redundant)된다. : 어떠한 편재 단백질이라도 기능적으로 동일한 형태는 극히 많은 수이다. (즉, 단백질 서열)
(P4) 어떠한 종에서도 특정 편재 유전자는 불필요하다. : 명백하게, 모든 유기체가 동일한 서열을 갖거나 혹은 비슷한 서열을 가져야 할 선험적인 근거는 없다. 어떠한 특정 유기체에는 기능적으로 필요한 특정 서열은 없으며, 오직 필요한 편재 유전자나 단백질은 수 없이 많은 기능적으로 동일한 유전자 형태 중에서 한 가지만 있으면 되는 것이다.
(P5) 기능을 위해서 유전과 서열이 서로 상관관계가 있을 필요가 없음에도 불구하고, 유전과 서열은 상관관계가 있다. : 두 개의 다른 유기체가 서로 유사한 서열을 가진 편재 단백질을 가질 수 있는 메카니즘은 이 세상에는 오직 단 한가지만이 관찰되며 그 메카니즘은 유전이다.
(C) 그러므로, 비슷한 편재 유전자는 genealogical relationship을 나타내는 것이다. : 서로 비슷한 편재 단백질의 서열을 가지고 있는 유기체는 서로 계보가 서로 관계가 있다는 결론을 내리게 한다.
토의:
DNA 염기 서열 분석 기술이 나타나기 전에는 단백질의 아미노산 서열이 종들 사이의 계통발생학적 관련성을 확립하는데 사용되었습니다. 기능을 가진 유전자의 서열 연구는 서로 편재(즉, 모든 유기체가 가지고 있는)하는 단백질의 유전자 (혹은 RNA)에 집중되었습니다. 이것은 전체 종들의 표현형과 무관하게 비교한다는 것을 확실히 하기 위해서입니다.
예를들어, 우리가 침팬지와 사람의 단백질 서열을 비교한다고 가정하는 경우, 사람과 침팬지 모두는 서로 비슷한 해부학적 특징과 기능을 많이 가지고 있습니다. 그러므로 우리는 설사 그들이 계보가 전혀 관련이 없을 지라도 그들의 단백질이 매우 비슷할 것이라고 기대할 수 있습니다. 그러나, 우리는 모든 살아있는 유기체가 사용하는 매우 기본적인 유전자 예를들어 사이토크롬 c 유전자를 비교할 수도 있으며 이 유전자는 침팬지와 사람의 특정한 특징에 어떠한 영향을 미치지 않습니다.
사이토크롬 c는 매우 필수적이고, 진핵생물과 박테리아를 포함해서 어느 유기체나 가지고 있는 편재 단백질입니다(Voet and Voet 1995, p. 24). 세포의 미토콘드리아는 사이토크롬 c를 가지고 있으며, 사이토크롬은 산화성 인산화(oxidative phosphorylation) 이라는 기본적인 대사과정에서 전자를 전달하는데 사용됩니다. 우리가 호흡하는 산소는 이 과정을 통해서 에너지를 발생하는데 사용되는 것입니다(Voet and Voet 1995, pp. 577-582).
사이토크롬 c와 같은 편재 유전자를 사용함으로, 두 유기체가 서로 계통학적으로 관련이 없음에도 불구하고 서로 동일한 단백질 서열이나 혹은 비슷한 단백질 서열을 가질 것이라고 가정할 이유가 없어졌습니다. 이것은 부분적으로는 단백질 서열의 기능 중복성 (functional redundancy)과 구조에 근거하는 것입니다. 여기서 "기능 중복성"은 수 많은 다른 단백질 서열이 동일한 일반적인 구조를 가지는 것과 동일한 일반적인 생물학적 역할을 수행한다는 것을 나타내는 것입니다. 사이토크롬 c 는 기능적으로 극히 중복될 수 있는 단백질일 것입니다. 왜냐하면 서로 다른 수 많은 서열이 사이토크롬 c 전자 전달 단백질을 형성하기 때문입니다. 서로 다른 단백질의 기능이 중복된다고 해서 수행능력이 정확하게 동일할 필요까지는 없습니다. 그중 일부의 사이토크롬 c 서열은 다른 것보다 전자 전달 능력이 우수할 수도 있으나, 이 논증의 목적에서 이것은 큰 문제가 아닙니다.
수 십년 간의 생화학적 증거들은 대부분의 아미노산 돌연변이 특히 표면에 노출된 아미노산 잔기는 단백질의 기능 혹은 단백질의 구조에 거의 영향을 미치지 않는다는 것을 보여왔습니다 (Harris et al. 1956; Li 1997, p. 2; Matthews 1996). 가장 놀라운 사례는 바로, 다양한 박테리아의 거의 서열의 유사성이 없는 c-type 사이토크롬입니다. 이들은 서로 서열이 다름에도 불구하고 모두 같은 3차원 구조로 접혀지고 모두 동일한 생물학적 역할을 수행합니다( Moore and Pettigrew 1990, pp. 161-223; Ptitsyn 1998).
같은 종 사이에도 대부분의 아미노산 돌연변이는 기능적으로 변화를 주지 않습니다. 예를 들어, 인간의 헤모글로빈은 약 250가지의 돌연변이가 알려졌으며, 이러한 유전자를 가진 사람은 전 인류의 3%가 넘으나, 이 유전자를 염색체 한쌍에 모두 가진 사람이나 혹은 염색체의 한쪽에만 가진 사람이나, 임상적인 이상 증상을 보이지 않습니다 (Bunn and Forget 1986; Voet and Voet 1995, p. 235). 이러한 단백질의 기능 중복은 매우 일반적이며, 알려진 모든 단백질과 유전자에게서 관찰됩니다.
이것을 염두에 두고 사이토크롬 c 분자 서열을 다시 한번 생각해 보겠습니다. 사이토크롬 c는 생명체에 필수적입니다. 유기체는 이것이 없으면 살 수 없습니다. 비록 인간과 효모는 단백질의 40%가 서로 다름에도 불구하고, 자신의 사이토크롬 c 유전자가 제거된 효모(단세포 생물의 하나)에서 인간의 사이토크롬 c가 제대로 작용했습니다(Tanaka et. al 1988a; Tanaka et al. 1988b; Wallace and Tanaka 1994). 사실상, 다랑어(어류), 비둘기(조류), 말(포유류), 초파리(곤충), 그리고 쥐(포유류)의 사이토크롬 c 는, 자체 사이토크롬 c가 제거된 효모에서 모두 기능을 했습니다(Clements et al. 1989; Hickey et al. 1991; Koshy et al. 1992; Scarpulla and Nye 1986). 게다가 사이토크롬 C 유전자 분석을 강도있게 실시한 결과, 단백질 서열의 대부분이 생체내에서 기능을 하는데 불필요하다는 것이 알려졌습니다(Hampsey et al. 1986; Hampsey et al. 1988). 사이토크롬 C 의 100개 아미노산 중에서 기능에 필요한 것은 오직 1/3뿐입니다. 사이토크롬 C 의 아미노산의 대부분은 hypervariable(즉, 비슷한 다른 많은 아미노산으로 대체가 됨) 합니다(Dickerson and Timkovich 1975). 더욱 중요한 것은 Hubert Yockey의 조심스러운 연구에서는 이러한 유전자 돌연변이 분석에 근거를 두고, 기능을 가진 사이토크롬 C 단백질 서열이 최소한 약 2.3 x 1093나 된다는 것을 계산하였습니다(Hampsey et al. 1986; Hampsey et al. 1988; Yockey 1992, Ch. 6, p. 254). 좀더 쉽게 말하면, 1093 이라는 숫자는 우리가 볼 수 있는 우주의 모든 원자의 수 보다 십억배나 더 큰 수입니다. 그러므로 기능을 가진 사이토크롬 C 서열은 거의 수적으로는 제한이 없으며, 서로 다른 두종이 서로 동일하거나 혹은 매우 비슷한 사이토크롬 C 단백질 서열을 가질 선험적인 이유는 없습니다.
과학적인 통계 분석의 용어로 말해서 "영가설(null hypothesis, 아무런 가정하지 않는 가설")은 인간과 침팬지의 사이토크롬 C 단백질의 비필수적인 아미노산의 성분은 서로 각각 무작위적일 것이라는 것입니다. 그러나, 공통 조상과 우리의 표준 계통발생수로부터 우리는 인간과 침팬지가 매우 가깝다는 것을 알고 있습니다. 우리는 그러므로, 확률에도 불구하고 단순히 유전 때문에, 인간과 침팬지의 사이토크롬 C 서열은, 예를 들어 인간과 효모의 사이토크롬 C 보다 훨씬 더 가까울 것이라고 예측할 수 있습니다.
확인 :
인간과 침팬지는 정확히 동일한 사이토크롬 C 단백질 서열을 가지고 있습니다. 위에 주어진 "영가설"은 거짓입니다. 공통 조상이 없다면, 이러한 것이 일어날 확률은 아무리 많아야 10-93(1093중에 하나)입니다. 그러므로 우리는 단백질의 이러한 고도의 유사성은 공통 조상 이론의 놀라운 보강 증거입니다. 게다가, 인간과 침팬지의 사이토크롬 C 단백질은 다른 포유류의 것과 비교해서도 10개 정도 차이가 날 뿐입니다. 유전과 같은 메카니즘이 없다면 이러한 것이 일어날 확률은 10-29에 불과합니다. 효모인 칸디다 크루세이(Candida krusei)는 인간으로부터 가장 먼 친척관계에 있는 진핵 생물의 하나입니다. 칸디다는 인간의 서열과 51개 아미노산의 차이를 보이며, 아무리 높게 잡아도 이 확률은 10-25이하입니다.
한가지 가능한 그러나 별로 가능성은 없는 반대 주장은 다양한 사이토크롬 C 의 기능수행력의 약간의 차이가 이러한 서열의 유사성을 가지고 왔다는 것입니다. 이 반대 주장은 수행능력이 거의 표현형적으로 구분할 수 거의 동일한 서열이 수도 없이 많다는 것을 생각하면 별로 사실일 것 같지 않습니다. 게다가, 거의 동일한 서열이라고 해서 이것의 수행 능력이 거의 비슷한 것은아닙니다.
그럼에도 불구하고, 논의를 계속하기 위해서, 활발한 대사 혹은 빠른 속도의 근육 수축을 하는 유기체에서 전자를 좀 더 빨리 전달하는 사이토크롬 C가 필요하다고 가정할 수 있습니다. 만약 이것이 사실이라면, 우리는 비슷한 환경이나 비슷한 생리학적 요구와 이러한 서열의 유사성은 어떠한 패턴을 보일 것이라고 기대할 수 있습니다. 그러나 그러한 것은 관찰되지 않았습니다. 예를들어, 박쥐의 사이토크롬 C는 벌새의 사이토크롬 C 보다는 인간의 사이토크롬 C와 더 유사했습니다. 돌고래의 사이토크롬 C는 상어의 사이토크롬 C 보다는 인간의 사이토크롬 C와 더 비슷했습니다. 앞의 예측 1.3에서 언급했듯이, 사이토크롬 C 자료로부터 구성된 계통발생수는, 완전히 독립적인 자료인 형태학적 자료로부터 결정된 주료 생물군(taxa)의 친척관계와 정확하게 되풀이 되었습니다(McLaughlin and Dayhoff 1973). 이러한 사실은 또한 사이토크롬 C 서열이 표현형적인 기능에 독립적이라는 것을 추가적으로 지지할 뿐이었습니다. (전자를 전달할 때 분명히 기능이 살아있는 사이토크롬 C가 필요하다는 것을 제외하면)
반복:
이 예측은 예측 1.3 "서로 독립적인 계통발생수의 일치"와는 약간 차이가 있습니다. 위에 제시한 증거들은 많은 편재 단백질(사이토크롬 c 와 같은)에 관한 것이며, 어떤 한 유기체에 존재하는 단백질을 만들 수 있는 동등한 서열은 매우 많다는 것입니다. 우리가 두 유기체에서 편재 단백질에 대해서 동일하거나 매우 비슷한 서열을 발견할 때마다 뭔가 이상한 것이 진행되고 있다는 것을 알 수 있습니다. 왜 두 유기체는 반대로 일어날 가능성이 천문학적으로 높은데도 불구하고 서로 비슷한 편재 단백질을 가지게 된 것인가? 우리는 기능적으로 그러할 필요가 없음에도 불구하고 서로 비슷한 단백질 서열을 갖는 것에 대해서는 오직 한가지 이유밖에는 알지 못합니다. 그리고 그것은 바로 유전입니다. 그러므로, 이러한 경우에 우리는 확실하게 두 유기체의 계보가 관련이 있다고 연역 추론할 수 있습니다. 이러한 의미에서 서열의 유사성은 공통 조상 이론의 테스트일 뿐만 아니라, 공통 조상은 유전과 서열의 유사성에 대한 관찰로부터 연역적인 결과이기도 합니다. 마지막으로, 사이토크롬 C에서 관찰된 유사성은 이 편재 단백질 하나에만 해당되는 것이 아닙니다. 침팬지와 인간사이의 모든 비교된 편재 단백질이 서로 비슷했으며, 비교된 것은 많이 있었습니다.
반증가능성:
공통조상 이론을 가정하지 않으면, 가장 그럴듯한 결과는 서로 다른 유기체간의 사이토크롬 C 단백질 서열은 서로 매우 다를 것입니다. 만약 그랬다면, 계통발생 분석은 불가능했을 것이고, 계통적으로 친척관계가 없었을 것이며, 동시에 종의 기원과 관련이 없다는 강력한 증거가 되었을 것입니다(Dickerson 1972; Yockey 1992; Li 1997).
게다가, 이 논증의 기초는 만약 (1) 한 종에 특이적인 사이토크롬 C 단백질은 오직 그 유기체에서만 독점적으로 기능한다. 혹은 (2) 자신이 가진 본연의 한 사이토크롬 C와 같은 기능을 하는 다른 사이토크롬 C 서열은 없다. (3) 유전 말고 다른 관찰이 된 그 어떠한 메카니즘이 편재 단백질과 유기체의 특정 형태와 인과적으로 상관관계를 맺어준다는 것이 증명되면 아주 쉽게 손상되었을 것입니다.
단백질 서열의 유사성처럼, 두 종의 편재 유전자의 DNA 서열의 유사성도 공통 조상을 암시합니다. 물론, 사이토크롬 C와 같이 conserved 한 단백질의 종합적인 DNA 서열 비교는 간접적으로 아미노산 서열에 대해서 고려해야 합니다. 왜냐하면 DNA 서열이라는 것은 단백질의 서열을 표현하는 것이기 때문입니다. 그러나, DNA 서열 자체도 또 다른 수준의 중복(redundancy)가 있습니다. 유전자 코드 자체가 정보적으로 중복되어 있습니다. 평균적으로 3개의 다른 코돈(DNA 염기 세 개를 코돈이라고 한다)이 같은 아미노산을 나타냅니다(Voet and Voet 1995, p. 966). 그러므로 사이토크롬 C에 대해서는 동일한 단백질 서열에 대해서도 대략 3104, 즉 1046가 넘는 DNA 서열이 있을 수 있습니다.
우리는 우리의 예측을 매우 자세하게 할 수 있습니다. 2가지의 기능을가진 사이토크롬 C 유전자의 어떠한 염기 서열의 차이도 필연적으로 기능에는 차이가 없거나 거의 차이가 없습니다. 인간(그리고 다른 대부분의 포유류에 있어서)에게 있어서 돌연변이 비율은 한 세대당 한 곳에서의 비율은 ~1-5 x 10-8로 측정되고 있으며, 평균적인 영장류의 한 세대는 20년입니다(Mohrenweiser 1994, pp. 128-129). 화석 기록으로부터 우리는 인간과 침팬지가 공통조상으로부터 갈라져 나온 것은 1천만년전이 안 된다는 것을 알고 있습니다(넉넉하게 계산해도 600만년 전이다.)(Stewart and Disotell 1998). 그러므로, 만약 침팬지와 인간이 진정으로 가계가 서로 관련이 있다면, 우리는 이들 각각의 사이토크롬 C 유전자의 DNA 서열이 3% 이내라고 예상할 수 있습니다. - 사이토크롬 C 유전자의 필수 기능 때문에 아마도 이보다 훨씬 적을 것입니다. -
확인:
위에서 언급한 것과 마찬가지로, 침팬지와 인간의 사이토크롬 C 단백질은 동일합니다. 그리고 결정적으로 인간과 챔팬치의 사이토크롬 C DNA 서열은 이 단백질을 만들 수 있는 염기 서열이 1049가지나 있음에도 오직 염기 하나만 (0.3% 차이) 다를 뿐입니다.
DNA 코딩 중복(redundancy)과 단백질 서열의 중복성이 합쳐진 효과는 DNA 서열 비교가 2가지의 중복성을 갖게 합니다. 편재 단백질의 DNA 서열은 표현형과는 상관관계가 전혀 없습니다. 그러나 그들은 유전에 의해서 인과적으로 상관관계를 갖게 된다. 이것이 계통학에서 DNA 서열이 매우 튼튼한 것으로 간주되는 이유입니다.
반증가능성:
가장 그럴듯한 결과는 이러한 단백질을 코딩하는 DNA 서열이 매우 심하게 다른 경우입니다. 이렇게 된다면 대진화에 대한 완전한 반증이 될 것이며, 침팬지와 인간의 계통이 전혀 관련되어 있지 않다는 매우 강력한 증거가 될 것입니다. 물론, 예측, 4.1의 반증가능성도 DNA 서열에 그대로 적용시킬 수 있습니다.
트랜스포존은 바이러스와 매우 유사합니다. 그러나, 그들은 바이러스 외피 단백질이 없으며, 세포 경계를 넘어갈 수 없기 때문에 그들은 오직 그들의 숙주 게놈에서만 복제를 할 뿐입니다. 그들은 게놈내부의 기생물(parasites)로 생각될 수 있습니다. 극히 예외적인 상황을 제외하고는 그들이 한 후생동물에서 다른 것으로 전달되는 유일한 방법은 직접적인 DNA 복제와 유전에 의한 것 뿐입니다. (즉, 당신의 트랜스포존은 당신의 자녀에게 전달됩니다.) (Li 1997, pp. 338-345)
트랜스포존의 복제는 자신을 스스로 복제해서 복제된 DNA를 숙주의 게놈에 무작위로 다른 곳에 끼워넣는 것을 의미합니다. 트랜스포존 복제(transposition이라고도 불림)은 효모, 옥수수, 왈라비, 인간, 박테리아, 그리고 파리를 포함한 많은 유기체에서 직접적으로 관찰되었으며, 최근에 이 메카니즘이 아주 잘 이해되었습니다. (Li 1997, pp. 335-338;Futuyma 1998, pp. 639-641). 레트로트랜스포지션의 특수한 관찰사례에서 이것이 인간에서 신경섬유종 증(多發神經纖維腫症) 과 혈우병을 일으키는 것으로 알려졌습니다(Kazazian et al. 1988; Wallace et al. 1991).
서로 다른 두 종에서 동일한 염색체 위치에서 동일한 트랜스포존을 발견하는 것은 공통조상에 대한 강력하고 직접적인 증거가 됩니다. 왜냐하면 그들은 무작위적으로 끼어들어가고 일반적으로 유전이 아닌 다른 방법으로는 이것이 전달되지 않기 때문입니다. 게다가, 한번 이 공통조상이 트랜스포지션이 일어났다고 추론되는 경우, 이 공통조상의 모든 후손들이 같은 트랜스포지션을 갖게 됩니다. 가능한 예외에는 결실이 일어나서 이 트랜스포지션이 제거되는 경우입니다. 그러나 결실은 완전히 깨끗이 제거하는 경우는 없으며, 일반적으로 트랜스포존의 서열 일부를 남기고 있습니다.
확인:
일반적인 트랜스포존의 한가지는 SINE retroelement 입니다(Li 1997, pp. 349-352). 그리고 중요한 SINE transposon의 하나는 300 bp Alu element입니다. 모든 포유류는 많은 Alu elements를 가지고 있으며 인간을 게놈의 10%가 이것으로 구성되어 있습니다.(즉, 반복되는 유전자가 모두 6천만 염기에 이름니다.)(Smit 1996; Li 1997, pp. 354, 357). 매우 최근 인간의 Alu transpositions 은 인간의 역사이후 와 선사시대의 인간의 이주에 대해서 밝히는데 사용되었습니다. 왜냐하면, 일부 사람들은 다른 사람들이 가지고 있지 않은 새로운 Alu insertion을 가지고 있기 때문입니다(Novick et al. 1993; Novick et al. 1995). 실제로, 일반적인 Alu 트랜스포지션은 친자소송이나 범죄 과학수사에서 공통 조상의 신뢰할 만한 표지라는 것이 잘 보여졌습니다(Novick et al. 1993; Novick et al. 1995; Roy-Engel et al. 2001). 매우 중요하게도, 사람의 ?-globin 클러스터는 7개의 Alu 엘리멘트가 존재하며, 이들 각각은 침팬지에서도 동일한 7군데에서 발견되고 있습니다(Sawada et al. 1985).
좀더 자세히 말하면, 3개의 서로 다른 SINE 트랜스포지션이 고래류, 하마, 그리고 반추동물에서 동일한 염색체 위치에서 발견되며 이들은 모두 표준 계통발생수에 따르면 서로 가깝게 관련되어 있습니다. 그러나, 낙타와 돼지를 포함한 다른 포유류는 이 3가지의 특정 트랜스포지션이 없습니다(Shimamura 1997).
좀더 자세한 내용과 설명은 에드워드 막스의 Plagiarized Errors and Molecular Genetics FAQ에 잘 나와 있습니다.
기능이 없으면서 공통조상의 증거를 제공하는 분자의 다른 예는 가유전자입니다. 그들이 조절유전자가 문제가 있거나, 혹은 발현되지 못하도록 내부에 종료 서열이 있는 경우를 제외하고는 가유전자는, 기능을 가진 자신들의 counterparts (일차 서열와 종종 염색체에서의 위치)와 매우 가깝게 관련되어 있습니다. 그것들은 기능이 없으며 제거되도 유기체의 표현형에는 영향을 미치지 않습니다. 가유전자는 만약 이것이 어떠한 것의 흔적이 아니라면(예측 2.2.와 같은 사례), 유전자 복제(역주 : 여기서 말하는 복제는 정상적인 세포 분열에 의해서 복제되는 것이 아님)에 의해서 만들어진 다음에 돌연변이 된 것입니다. 트랜스포지션, 염색체 복제, 그리고 염색체의 불균등 교차(unequal crossing over)와 같이 유전자 복제를 일으킬 수 있는 많은 과정들이 관찰되었습니다. 트랜스포지션(예측 4.3 비교)와 같이 유전자 복제는 매우 드믈면서도 무작위적인 사건이며, 물론, 복제된 DNA는 모두 유전됩니다. 그러므로 두 종 사이에서 동일한 가유전자를 동일한 염색체 위치에서 발견하는 것은 공통조상에 대한 강력한 증거가 됩니다.
확인:
영장류와 인간사이에 가유전자를 공유하고 있는 많은 사례가 있습니다. 그중 하나가 ψη-globin으로 이것은 일종의 헤모글로빈 가유전자입니다. 이것은 영장류사이에서만 공유되고 있으며, 동일한 염색체 위치에서 동일한 돌연변이가 일어나서 이 유전자가 기능을 하지 못하게 되었습니다(Goodman et al. 1989). 또 다른 사례는 스테로이드 21-하이드록실레이즈 유전자입니다. 인간은 스테로이드 21-하이드록실레이즈의 유전자를 2 copy 가지고 있으며 그중 하나는 기능이 있지만 하나는 기능이 없는 가유전자입니다. 기능이 있는 유전자를 비활성화시키면 '선천성 부신 과형성증(congenital adrenal hyperplasia, CAH)'이라는 매우 희귀하고 심각한 유전 질병이 생기게 됩니다. 침팬지와 인간은 모두 이 가유전자에서 동일한 8 bp 결실(deletion)이 생겨서 기능을 못하는 것입니다(Kawaguchi et al. 1992). 이 사례에서 가유전자의 비기능성은 positively demonstrated 되었다는 것을 강조합니다.
There are very many examples of shared pseudogenes between primates and humans. One is the ψη-globin gene, a hemoglobin pseudogene. It is shared among the primates only, in the exact chromosomal location, with the same mutations that render it nonfunctional (Goodman et al. 1989). Another example is the steroid 21-hydroxylase gene. Humans have two copies of the steroid 21-hydroxylase gene, a functional one and a nonfunctional pseudogene. Inactivation of the functional gene leads to congenital adrenal hyperplasia (CAH), a rare and serious genetic disease. Chimps and humans both share the same eight bp deletion in this pseudogene that renders it nonfunctional (Kawaguchi et al. 1992). Note that in this case, the nonfunctionality of the pseudogene has been positively demonstrated.
예측 4.5: 분자 증거 - 내재성 레트로바이러스
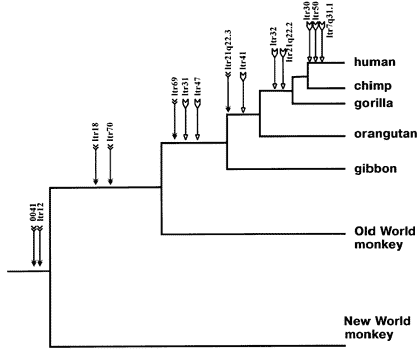
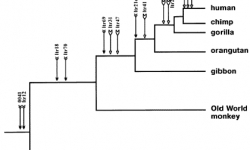
그림 4.4.1. 다양한 영장류에서의 동일한 염색체 위치에서의 인간 내재 레트로바이러스 K (HERV-K) 삽입(Reprinted from Lebedev et al. 2000, ? 2000, with permission from Elsevier Science). |
그리고 다른 사례는 내생 레트로바이러스(endogenous retroviruses)입니다. 내생 레트로바이러스는 과거의 기생적인 바이러스 감염의 분자적인 잔류물입니다. 종종, 레트로바이러스의 게놈의 복사본(copy)들이 숙주의 게놈에서 발견되며, 이러한 레트로바이러스 유전자 복제본들은 내생 레트로바이러스 서열들이라고 불립니다. 레트로바이러스(AIDS 바이러스 혹은 백혈병을 일으키는 HTLV1 와 같은 것)는 그들 자신의 바이러스 게놈의 DNA 복사본을 만들어 숙주의 게놈에 삽입시킵니다. (역주, 레트로 바이러스는 RNA 바이러스이며, 유전정보를 DNA로 복사해서 숙주의 게놈에 삽입시킵니다.) 만약 이러한 것이 생식세포 (예를들어 정자나 난자)에서 일어난다면 레트로바이러스 DNA는 숙주의 후손에 유전됩니다. 다시 말하지만, 이러한 과정은 매우 드믈고 거의 무작위적입니다. 그렇기 때문에 서로 다른 두 종에서 동일한 염색체 위치에서 레트로유전자를 발견하는 것은 공통조상을 나타내는 것입니다.
확인:
인간에 있어서, 내생 레트로바이러스는 전체 게놈의 1%에 해당하며, 전제 다 합하면 약 30,000개 가량의 서로 다른 레트로바이러스가 각 개인의 게놈 DNA에 박혀 있습니다(Sverdlov 2000). 최소한 7개의 서로 다른 공통 레트로진 삽입 사례가 침팬지와 인간 사이에 밝혀졌으며, 이 숫자는 두 유기체의 게놈 서열이 알려지면서 더 증가하고 있습니다(Bonner et al. 1982; Dangel et al. 1995; Svensson et al. 1995; Kjellman et al. 1999; Lebedev et al. 2000;Sverdlov 2000). 그림 4.4.1은 인간을 포함한 몇몇 영장류들의 계통발생수로, 이들 영장류들의 게놈에서 많은 공유 내생 레트로바이러스를 밝혀낸 최근 연구 결과로부터 얻은 것입니다. (Lebedev et al. 2000). 화살표는 숙주로 바이러스 DNA가 삽입된 시간을 상대적으로 나타낸 것입니다. 삽입점 이후의 모든 오른쪽으로 향하는 가지는 이들 레트로바이러스 DNA를 가지고 있습니다. 이것은 한번 어떤 유기체의 생식세포에 삽입되면 그로부터 유전된다는 사실을 반영하고 있습니다.
고양이과(예, 고양이)는 또 다른 사례를 제공합니다. 표준 계통발생수는 작은 고양이과 동물들이 큰 고양이과 동물보다 나중에 갈라져 나왔다는 것을 보여줍니다. 이 작은 고양이들(즉, 정글살쾡이, 유럽 야생고양이, 아프리카 야생고양이, 검은발 고양이, 그리고 집고양이)는 특정한 테르로바이러스 유전자 삽입을 공유합니다. 하지만 조사한 다른 육식동물들은 모두 이러한 레트로유전자가 없었습니다(Futuyma 1998, pp. 293-294).
반증가능성:
만약, 어떤 다른 포유류(예를들어, 개, 소, 오리너구리 등)이 동일한 염색체 위치에 동일한 레트로유전자를 가지고 있다면 대진화론적으로 말이 않되는 것입니다. 또 다른 사례로, 만약 개가 그림 4.4.1에서 보는 것과 같이 인간만이 가지고 있는 것으로 알려진 3가지의 HERV-K 삽입을 가지고 있는 다는 것은 극히 가능성이 없어 보입니다. 왜나하면 다른 어떠한 영장류도 이 레트로바이러스 서열을 가지고 있지 않기 때문입니다.
 진화론적인 기회주의
진화론적인 기회주의