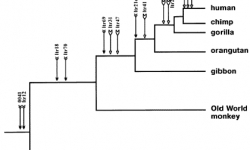
?표준 계통발생수를 연구함으로서, 모든 종들이 그 종만의 계통적인 역사가 있다는 것을 볼 수 있게 됩니다. 각 종은 original common ancestor과 연결되는 그들의 공통조상을 가지고 있습니다. 우리는 이 역사의 증거와 계보를 가지고 있는 유기체를 기대할 수 있습니다. 표준 계통발생수는 주어진 종에 대해서 어떠한 역사적인 증거가 가능하며 어떠한 것이 불가능한지 예측해 줄 수 있습니다.

|
대진화 역사를 통해서 특정기능은 생기기도 하고 사라지기도 합니다. 그러므로 우리는공통조상과 점진주의에 의한 제한 때문에 많은 생명체들이, 잃어 버린 기능의 구조적인 흔적인 흔적기관을 가지고 있을 것이라고 예측할 수 있습니다. (정확한 진화론적 메카니즘은 그것이 점진적인 한 여기에서는 필요하지 않습니다.) 이러한 것이 존재할 명백한 이유가 없기 때문에, 기능이 없는 특성들은 생명체의 특히 매우 혼동스럽게 하는 특징입니다. 또한 흔적 기관들은 다른 생명체의 동일하거나 좀더 발달된 구조와 비교할 경우, 서로 다르거나 혹은 상대적으로 경미한 기능을 가지고 있습니다. 확인: 생명체의 거의 쓸모 없거나 기능이 없는 특성에 관한 사례는 상당히 많으며, 이것들은 매우 자주 진화 역사로 설명됩니다. 예를들어, 뱀은 4다리가 달린 파출류의 후손이라고 알려져 있습니다. 대부분의 비단뱀(다리가 없는 뱀)은 그들의 피부아래 흔적기관적인 골판이 숨겨져 있습니다. (Cohn 2001; Cohn and Tickle 1999). 이 비단뱀의 흔적기관적인 골반은 척추에 붙어있지 않으며(대부분의 척추동물은 척추에 붙어있습니다.), 단지 복강에서 떠다닙니다. 일부 도마뱀은 그들의 피부아래 밖에서는 보이지 않는 흔적기관적이고 기능이 없는 다리를 가지고 있습니다(Raynaud and Kan 1992). 동굴에 살고 있는 많은 동물들, 예를들어 물고기 Astyanax mexicanus(멕시칸 테트라, 역주 : 국내에서는 '블라인드 케이브 카라신'이라고 불리며 구입이 가능합니다.)와 살라만더 종인 Typhlotriton spelaeus와 Proteus anguinus은 앞을 볼 수 없지만, 아직도 흔적기관적인 눈을 가지고 있습니다(Besharse and Brandon 1976; Durand et al. 1993; Jeffery 2001; Kos et al. 2001). 멕시칸 테트라의 눈은 앞을 볼 수도 없으며, 이러한 것이 필요 없음에도 불구하고, 렌즈를 포함하여, 퇴화한 망막, 퇴화한 시신경과 눈의 공막을 가지고 있습니다 (Jeffery 2001). 앞을 못보는 살라만더는 망막과 렌즈가 있는 눈을 가지고 있으나, 눈꺼풀이 눈위로 자라서, 외부의 빛을 차단하고 있습니다(Durand et al. 1993; Kos et al. 2001). 민들레는 reproduction 없이 reproduce 합니다(단위생식으로 알려졌습니다.), 그러나, 그들은 아직도 꽃과 꽃가루(이 두가지는 유성생식의 수정에 사용되는 기관입니다.)를 가지고 있습니다(Mes et al. 2002). 꽃과 꽃가루는 그러므로, 민들레에게는 쓸데 없는 특성인 것입니다. 인간의 조상은 식물성 물질을 씹고 가는데 첫째 어금니와 둘째 어금니가 필요한 것으로 알려져 있습니다.모든 성인의 90% 이상은 3번째 어금니(보통은 사랑니라고 알려졌습니다.)가 나옵니다. 일반적으로 이들 이빨은 잇몸에서 부터 나오는 않으며, 모든 개인중 1/3은 사랑니가 기형이거나 눌려져 있습니다(Hattab et al. 1995; Schersten et al. 1989). 이 쓸 데 없는 기능은 심한 통증을 일으키거나, 상처가 생길 위험이 증가시키거나, 혹은 병이 걸리거나 죽음에 이르게 할 수도 있습니다(Litonjua 1996; Obiechina et al. 2001; Rakprasitkul 2001; Tevepaugh and Dodson 1995). 융합된 날개 뚜껑아 완벽하게 형성된 날개를 감추고 있는, 날지 못하는 딱정벌레(예를들어 사슴벌레과의 바구미)도 많은 종류가 있습니다. 이들 모든 사례는 유기체의 예상 선조들에게 유익이 되었던 기능과 구조로 설명할 수 있습니다 (Futuyma 1998, pp. 122-123). 반증가능성: 어떠한 유기체도 그들의 선조중의 하나가, 기능을 가지고 있던 구조가 아닌 것을 흔적구조로 가질 수 없습니다. 그러므로 각각의 종에 대해서 표준계통발생수는 기능한 흔적이관과 가능하지 않은 흔적기관에 대해서 엄청난 수의 예측을 할 수 있습니다. 흔적기관이 아니라, 공유하고 있는 파생 특성과 분자생물학적 유전자 서열 자료로 계통발생수 및 예상 공통조상의 특성을 결정할 수 있습니다. 그러므로 만약 공통조상이 거짓이라면 흔적 기관은 아마도 진화론적으로 설명할 수 없을 것입니다. 예를들어 고래는 유선, 태반, 아래턱의 뼈가 하나인 것 등등의 많은 기준에 의해서 포유류로 분류됩니다. 뱀도 이와 비슷하게 여러 가지 파생 특징으로 인하여 파충류로 분류됩니다. 그러나 이론적으로는 뱀이나 고래가 어류로 분류될 수 있을 수도 있습니다. 만약 그렇다면 흔적기관으로서 다리나 뱀의 흔적기관적인 골반은 진화론적으로 말도 않되는 것이며 공통조상 주장과 일관성이 없어집니다. 우리는 어떠한 양서류, 조류, 파충류에서도 태반이나 흔적기관적인 젖꼭지를 발견하지 못할 것입니다. 어떠한 포유류도 흔적기관적인 날개를 발견할 수 없을 것입니다(하지만 그들은 흔적기관 꼬리를 가질 수 있습니다. 인간처럼). 우리는 흔적기관 척추를 가진 곤충을 발견하지 못할 것입니다. 이 예측은, 흔적기관이라고 생각되었던 구조가 어떠한 기능을 가지고 있다는 것으로 부정되지 않습니다. 이러한 현상이 일어나면 이 구조는 단지 예측 3.1에서 다루는 상동기관이 되는 것이거나, 혹은 예측 3.5에서 다루는 불충분한 설계의 사례가 될 것입니다. 일부 반진화론 저자들은 과학적으로나 신학적으로 어떠한 구조가 기능이 전혀 없다는 것을 보이는 것이 불가능하다고 합니다 (e.g., see Ham et al. 1990; Scadding 1981). 이러한 허무주의적인 철학은 실용적인 과학에는 설자리가 없습니다. 이러한 잘못된 결론은 가설을 검증하는데 부정적인 증거가 사용될 수 없다는 잘못된 전제에 근거하고 있는 것입니다. 그러나, 부정적인 증거는 만약 적절한 실험적인 대조구가 있었다면 받아들여지는 것입니다. 물리학에서 일어날 수 있는 이것과 비슷한 다음과 같은 것을 생각하면 부정적인 증거도 분명히 쓸모가 있는 것입니다. 만약 어떤 구조가 기능이 전혀 없다는 것을 증명하는 것이 불가능하다면, 같은 이유로 주어진 어떤 원소가 방사능이 없다는 것을 증명하는 것이 불가능할 것입니다. 그러나 물리학에서는 납-207이 방사능이 없다는 것은 아주 잘 확립되었습니다. 이것은 예를들어 납-207에서는 방사능이 검지되지 않지만, 인-32와 같은 다른 원소에서 방사능을 검지할 수 있기 때문입니다. 물리학에서 인-32는 양성대조구이며 이것은 납-207에서 얻은 negative evidence에서 사용하기 위해 필요합니다. 마찬가지로 주어진 구조가 기능이 없다는 것은, 이와 동시에 같은 환경에서 분명히 유용하게 보이는 다른 종의 구조에서 유익한 기능을 발견할 수 있을 때 밝힐 수 있습니다. | |
Figure 2.1.1. Various organisms displaying nonfunctional, vestigial characters. From top to bottom: A. A blind cave salamander - look closely for the eyes buried underneath the skin. B. Astyanax mexicanus, the Mexican tetra, a blind cave fish. C. Apterocyclus honolulensis, a flightless weevil. The black wing covers cannot open, as they are fused, yet underneath are perfectly formed beetle wings. D. The nonfunctional flower of Taraxacum officinale, the common dandelion. E. A nonfunctional pollen grain from the dandelion. |
예측 2.2: 격세유전 (Atavisms)
해부학적 격세유전은 흔적기관과 개념적으로 매우 관련되어 있습니다. 진화론적으로 먼 조상은 가지고 있었으나, 가까운 조상에서는 발견되지 않는 잃어 버린 특징이 다시 나타나는 것을 격세유전이라고 합니다. 격세유전에는 다음과 같은 몇가지 기본적인 요소가 있습니다. (1) 성인기에 존재한다. 2) 부모나 가까운 조상에서는 보이지 않는다. 3) 집단내에서 매우 희귀하다(Hall 1984). 물론 진화론적으로 바라보지 않는다면 우리는 격세유전이 먼 조상이 가지고 있었으나 최근 조상은 잃어 버린 것이라고 말 할 수는 없습니다. 그러므로 우리는 여기에 주로 격세유전적인 구조의 가능성이 있는 것, 즉 자신이 속한 생물군(taxa)에서 볼 수 없는 특징을 가진 것만을 주로 관심을 가질 것입니다. 가상적인 사례로서 만약 돌연변이 말이 우연히 아가미를 가진다면 이것은 격세유전의 가능성이 있습니다. 왜냐하면, 아가미는 말이 속한 생물군의 특징이 아니기 때문입니다. 발생학적 이유로, 종종 공통조상을 가진 후손들에게서 가끔 격세유전이 나타나는 것으로 예측됩니다(Hall 1984; Hall 1995).
흔적기관과 마찬가지로, 만약 구조나 기능이 조상과 후손 계보중간에 사려졌다면 어떠한 유기체도 조상이 가진 적이 없는 격세유전적인 구조를 가질 수 없습니다. 그러므로 각 종에 대해서 표준계통발생수는 어떤 주어진 종에 대햇 엄청나게 많은 예상 격세유전과 불가능한 격세유전을 예측할 수 있습니다.
확인:
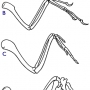
가장 잘 알려진 격세유전은 고래에서 발견됩니다. 표준계통발생수에서는 고래는 뒷발을 가진 육상 포유류에서 유래했습니다. 그러므로 우리는 고래의 돌연변이중에서 아주 드믈 게 뒷다리를 가진 것이 있을 수 있다고 기대할 수 있습니다. 사실상, 야생의 고래에게서 흔적기관적이고 격세유전적인 뒷다리를 가진 많은 사례가 발견된 사례가 있습니다(for reviews see Berzin 1972, pp. 65-67 and Hall 1984, pp. 90-93). 뒷발은 수염고래(Sleptsov 1939), 혹등고래(Andrews 1921) 및 많은 향유고래(Abel 1908; Berzin 1972, p. 66;Nemoto 1963; Ogawa and Kamiya 1957; Zembskii and Berzin 1961) 종에서 발견되었습니다. 대부분의 사례에서 이 고래들은 대퇴골, 정강이뼈, 및 비골(fibulae)을 가지고 있었지만 일부는 완전한 발가락을 가진 발이 있기도 했습니다.
격세유전에는 (1)드믈게, 말에서 고대의 메조히푸스나 메리치푸스에서 볼 수 있는 2번째 혹은 4번째 발가락이 형성되는 것, (2) 연작류새와 참새에서 나타나는 격세유전적인 넓적다리 근육, (3)개의 설골 근육, (4) 집게벌래의 날개(보통은 날개가 없음) (5) 새의 격세유전적인 비골(비골은 정상적으로는 극히 작음), (6) 기니픽과 살라만더의 여분의 발가락 및 (7)인간의 다양한 격세유전적인 특징(그중 하나를 다음에 자세히 설명할 것입니다.)를 포함한 다양한 사례들이 있습니다(Hall 1984).
의학적인 관심 때문에 인간은 가장 특성이 잘 알려져 있으며 많은 발생상의 비정상 현상이 알려졌습니다. 다른 포유류와 함께 공통 유전적인 유산을 반영하는 인간의 몇몇 격세유전이 있습니다. 가장 놀라운 것 중의 하나는 매우 드믄 "진짜 인간 꼬리(일반적으로 "미골 과정", "미골 돌출", "꼬리 덧돌기", "흔적기관 꼬리"라고 불립니다.)의 존재입니다. 100건 이상의 인간꼬리가 의학 문헌에 잘 보고되었습니다. 잘 문서화된 사례중에 의학적으로 "가짜-꼬리"로 알려진 것은 1/3 이하입니다(Dao and Netsky 1984; Dubrow et al. 1988). 가짜 꼬리는 진짜 꼬리가 아닙니다. 이들은 단지 꼬리쪽 부분에서 우연히 발견되는 다양한 형태의 단순한 병소에 불과할 뿐입니다. 종종 척주(脊柱), 미골(coccyx) 및 다양한 기형과 관련되어 있습니다. 이와 반대로, 진정한 격세유전적인 인간의 꼬리는 인간의 태아 발달과정에서 발견되는 배아의 꼬리의 가장 끝부분에서 발달되며(Figure 2.3.1 및, 정상적인 인간 배아의 꼬리에 관한 아래의 토론 ; Belzberg et al. 1991; Dao and Netsky 1984), 그리고 이것이 일반적으로 자연에서 큰 불이익이 없습니다(Dubrow et al. 1988; Spiegelmann et al. 1985). 진정한 인간의 꼬리는 지방조직, 연결조직, 중간에 길게 배열된 가로무늬근, 혈관, 신경섬유, 신경절 세포들, 및 특수화된 압력 감지 신경 기관의 복잡한 배열이 특징입니다. 이것에는 일반적으로 모낭이 충분하고, 피지선이 있는 피부로 덮혀 있습니다(Dao and Netsky 1984; Dubrow et al. 1988; Spiegelmann et al. 1985). 진정한 인간의 꼬리는 1인치에서 5인치 길이에 이르고(신생아) 이것을 움직이고, 다른 것에 접촉시킬 수 있습니다 (Baruchin et al. 1983; Dao and Netsky 1984; Lundberg et al. 1962). 비록 인간의 꼬리가 일반적으로 뼈대구조(일부 의학논문에서는 진정한 꼬리는 전혀 척추가 없다고 주장합니다.)가 없지만 몇몇 인간의 꼬리는 연골과 5개까지 되기도 하는 잘 발달되고, 분명한 척추뼈가 발견되기도 합니다(그림 2.2.1; Bar-Maor et al. 1980; Dao and Netsky 1984; Fara 1977; Sugamata et al. 1988). 그러나 미골 척추는 포유류 꼬리에 꼭 필요한 부분이 아닙니다. 의학문헌에 자주 보고되는 것과는 달리, 마카카 실바누스(Macca sylvanus) "바바리 유인원(Barbary ape)"의 2인치 길이의 흔적기관같은 꼬리에서 보듯이, 척추뼈가 없는 유인원의 꼬리 사례가 최소한 1건 이상 알려졌습니다(Hill 1974, p. 616; Hooten 1947, p. 23). 진정한 인간의 꼬리는 유전되는 경우가 극히 드믈지만 가족사에서 나타나는 경우가 있습니다(Dao and Netsky 1984; Ikpeze and Onuigbo 1999; Standfast 1992; Touraine 1955). 다른 격세유전적인 구조처럼, 일간의 꼬리는 somatic 혹은 germline의 돌연변이가 일어나서 인간 게놈에 보존되고 있는, 숨겨진 발생과정이 다시 활성을 얻었기 때문일 것입니다(Dao and Netsky 1984; Hall 1984; Hall 1995).
진정한 인간의 꼬리의 존재가 많은 종교적인 이유로 자극을 받은 반진화론자들 예를들어 듀안 기쉬 같은 사람들에겐 불행히도 상당한 충격이었다는 것을 지적할 필요가 있을 것 같습니다. 듀안기쉬는 자주 인용되는 "진화와 인간의 꼬리"라는 글을 썼습니다(Gish 1983; Menton 1994; ReMine 1982). 특정 단일사례만의 연구(Ledley 1982)에 근거하여 저자들은 엉터리로 격세유전적인 인간의 꼬리는 "단지 비정상적인 기형으로 어떤 상상의 조상의 상태로도 추적할 수 없다"고 잘못 결론 내렸습니다(Gish 1983). 그러나, 이들 논증은 가짜-꼬리에 대한 것이지, 진짜꼬리에 대한 것이 아닙니다. 왜냐하면, 진짜꼬리는, 근육, 혈관, , 척추, 및 연골의 복잡한 구조이며(Bar-Maor et al. 1980; Dao and Netsky 1984; Fara 1977; Sugamata et al. 1988), 이것은 움직이고, 접촉할 수 있으며, 종종 유전되기 때문입니다(Dao and Netsky 1984; Ikpeze and Onuigbo 1999; Standfast 1992; Touraine 1955). 게다가, 기쉬, 멘톤 및 레민 모두는 인간의 흔적기관 꼬리가 척추뼈가 없다면 진정한 꼬리가 아니라고 주장했으만, 이것은 앞서 마카카 실바누스가 척추뼈가 없는 살로만 이루어진 꼬리를 가졌다는 것을 생각하면 잘못된 주장입니다(Hill 1974, p. 616; Hooten 1947, p. 23).
반증가능성
앞의 흔적기관적인 구조와 기본적으로 동일합니다.
예측 2.3: 분자 수준의 흔적 특성들
흔적기관적인 특징은 분자수준에서도 발견될 수 있습니다. 인간은 아스코빅산(비타민 C)을 합성하는 능력이 없으며, 이 불행한 결과는 영향결핍이 일어나는 경우 괴혈병이 생기게 할 수 있습니다. 그러나 인간의 예상조상은 이 기능이 이었습니다. (영장류와 기니픽을 제외한 다른 대부분의 동물처럼) 그러므로 우리는 인간과 다른 영장류와 기니픽이 이 잃어 버린 기능에 대한 분자수준의 흔적 증거를 가지고 있을 것을 예측할 수 있습니다(nota bene, 이 예측은 니시키미와 다른 과학자들에 의해서 명시적으로 제시되었고, 아래에 설명한 연구를 하게 되는 원동력이 되었습니다.) (Nishikimi et al. 1992;Nishikimi et al. 1994).
확인:
최근에, 비타민 C 생합성에 필요한 유전자인 L-gulano-g-lactone oxidase 유전자가 인간과 기니픽에게서 발견되었습니다(Nishikimi et al. 1992;Nishikimi et al. 1994). 이것은 가유전자로 존재하며 유전자는 있지만 기능은 없었습니다(가유전자에 대한 것은 예측 4.4를 참조하시길 바랍니다). 사실 이글이 처음 쓰여진 이후로 비타민 C 유전자가 진화론에서 예측한 대로 다른 영장류에서도 발견되었습니다.우리는 이제 이 파괴된 유전자를 침팬지, 오랑우탄, 마칵 원숭이에게서 발견했습니다(Ohta and Nishikimi 1999). 그리고 예측대로, 기능이 없는 이 가유전자는 인간과 침팬지가 가장 비슷하고, 그 다음이 인간과 오랑우탄, 그리고 인긴과 마칵 원숭이가 가장 차이가 컷으며 이것은 진화론에서 예측한 대로 정확하게 맞는 것입니다. 게다가 이들에게 축적된 유전자의 돌연변이는, 가유전자와 같은 중성적인 DNA의 기본 돌연변이율에 대한 예측과 정확하게 일치했습니다(Ohta and Nishikimi 1999).
흔적기관적인 인간 유전자의 사례는, 다중 후각물질 수용체 유전자(Rouquier et al. 2000), RT6 단백질 유전자 (Haag et al. 1994), galatosyl transferase유전자(Galili and Swanson 1991)와 tyrosinase 관련 유전자(TYRL)(Oetting et al. 1993)을 포함한 여러 가지가 있습니다.
우리의 odorant receptor(OR)유전자는 현재는 잃어 버린 후각기능에 관련된 단백질의 정저를 저장했습니다. 우리의 예상 조상은 다른 포유류와 마찬가지로 우리가 현재 가진 것보다 더 민감한 후각기능을 가지고 있었습니다. 인간이 가지고 있는 99% 이상의 후각 수용체 유전자를 가지고 있으며, 이중 70%는 가유전자입니다. 생쥐나 마모셋과 같은 많은 다른 포유류는 우리가 가진 것과 동일한 OR 유전자를 가지고 있지만 이들 모두는 모두 작용합니다. 극단적인 사례는 육상동물의 후손인 돌고래로, 이들은 휘발성 odorant를 냄새 맡아야할 필요가 더 이상 없지만 아직은 많은 OR 유전자가 존재하며, 이들중 전부가 기능이 없습니다. 즉 이들 모두 가유전자입니다(Freitag et al. 1998).
RT6 단백질은 다른 오유류에서는 T 림프구의 표면에 발현됩니다만, 우리는 발현되지 않습니다. galatosyl transferase 유전자는 다른 동물의 세포막에서 발견되는 탄수화물을 만드는 것과 관련되어있습니다. tyrosinase는 모든 동물의 멜라닌 색소에 관련된 주된 효소입니다. TYRL은 tyrosinase의 가 유전자입니다.
우리는 이러한 유전자를 다른 영장류와 공유하고 있다는 것을 지적해야 할 것입니다. 그리고 이들 유전자를 기능이 상실되도록 만들었던 돌연변이도 공유합니다. (비기능성 특징을 공유하는 것에 대해서는 예측 4.3-4.5 참조하기 바랍니다.).
반증가능성:
만약 우리가 L-gulano-g-lactone oxidase 가유전자 혹은 다른 위에 언급한 흔적 유전자를 발견하지 못한다면 매우 혼란스러웠을 것입니다. 게다가 우리는 절대로 흔적 엽록체 유전자를 그 어떠한 후생동물 (예를들어 동물)에게서 발견하지 못할 것을 예측할 수 있습니다(Li 1997, pp. 284-286, 348-354).
예측 2.4: 개체발생과 유기체의 발생
![[Figure2.2.1 (atavistic tail)]](http://www.rathinker.co.kr/creationism/comdesc/images/tail.jpg) 그림 2.2.1. 6세 여아에게서 발견된 격세유전적인 꼬리의 X-선 영상 A radiogram of the sacral region of a six-year old girl with an atavistic tail. The tail was perfectly midline and protruded form the lower back as a soft appendage. The five normal sacral vertebrae are indicated in light blue and numbered; the three coccygeal tail vertebrae are indicated in light yellow. The entire coccyx (usually three or four tiny fused vertebrae) is normally the same size as the fifth sacral vertebrae. In this same study, the surgeons reported two other cases of an atavistic human tail, one with three tail vertebrae, one with five. All were benign, and only one was surgically "corrected" for cosmetic reasons (image reproduced from Bar-Maor et al. 1980, Figure 3.) |
![[Figure2.4.1 (cat embryo)]](http://www.rathinker.co.kr/creationism/comdesc/images/cat_embryo.jpg)
![[Figure2.4.1 (human
embryo)]](http://www.rathinker.co.kr/creationism/comdesc/images/human_embryo.jpg)
![[Figure2.4.2 (histological cross-section human of human embryonic tail at Carnegie stage 14)]](http://www.rathinker.co.kr/creationism/comdesc/images/tail_cross_section.jpg)

 진화론적인 기회주의
진화론적인 기회주의