진화론적인 기회주의(opportunism)는 진화론의 역사와 우발효과와 밀접하게 관련이 있습니다(일부 저자들은 기회주의 개념을 "연속성의 원리"라고 부릅니다.)(Crick 1968). 점진적인 변화를 가진 후손이라는 말은 새로운 유기체는 오직 그들이 전에 가지고 있었던 것만을 사용하거나 변화시킬 수 있다는 의미입니다. 새로운 구조와 기능은 그 전의 오래된 구조에서부터 새로 만들어져야 합니다(Futuyma 1998, pp. 110, 671-674). 이것은 기능과는 달리 구조는 엄격하게 유전되기 때문입니다. 진정한 그리고 전례없는 구조적인 새로움은 매우 드믄 것입니다. 이것은 진화의 가능한 경로에 대하여 상당한 제한을 가하고 있습니다.
예측 3.1: 해부학적 상동기관 (Anatomical paralogy)
점진주의로 인한 생명체 형태의 제한의 주요한 결과는 상동기관이 존재할 것이라는 예측입니다. 여기에서 쓰이는 용어 상동기관(Paralogy)은, 기능은 다르지만 구조가 매우 유사한 것을 말합니다. 한 종이 두 종으로 갈라질 때 한 종 혹은 두 종 모두 새로운 기능을 획득하게 됩니다. 새로운 종은 이 새로운 기능을 수행하기 위해서 기존의 구조를 새로 사용하거나, 변화시켜야 하기 때문에, 두 종에서 서로 같은 구조를 가진 것이 서로 다른 기능을 하게 됩니다. 이것을 상동기관이라고 합니다. 이것은 상동기관적인 구조는 한 가닥의 다른 진화론적인 증거로 설명할 수 있는 역사를 가지게 합니다. 이는 파생된 구조(새로운 기능과 구조)는 그보다 원시적인 (즉, 오래된) 구조에서 진화한 것이기 때문입니다. 결론적으로 중간화석의 가능한 형태에 대해서 자세하고 명백하게 드러나는 예측을 할 수 있습니다.
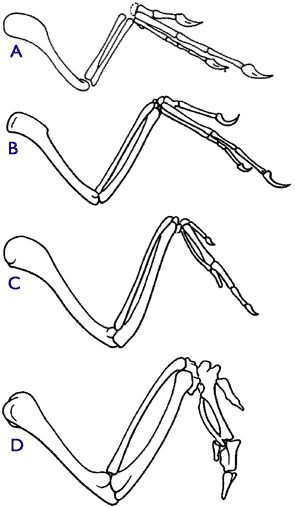
Figure 3.1.1. Comparison of the forelimbs of various relatives of modern birds. Forelimbs of (A)Ornitholestes, a theropod dinosaur, (B)Archaeopteryx, (C) Sinornis, an archaic bird from the lower Cretaceous, and (D) the wing of a modern chicken (modified from Carroll 1988, p. 340; Carroll 1997, p. 309). |
진화론적인 상동기관의 원리를 다시 말하면, 계통발생 수에서 예상되는 것은 기능적 연결성과는 달리, 구조적인 연결성을 가지고 있어야만 한다는 것입니다. 구조와 패턴은 유전되지만 기능은 유전되어야만 하는 것은 아닙니다. 조상으로부터 후손까지 한 계보를 따라가다 보면 기능은 생겼다가 사라질 수도 있지만 기반이 되는 구조는 하나에서 다른 것으로 점차적으로 변하며, 상대적으로 거의 변하지 않는 것으로 보입니다.
확인 :
현존하거나 멸종된 종들에게서 상동기관은 수도 없이 많습니다. 같은 뼈, 상대적으로 같은 위치에 있는 동일한 뼈가 원시적인 손, 박쥐의 날개, 새의 날개, 익룡의 날개, 고래와 펭귄의 지느러미 모양의 다리(flippers), 말의 다리, 두더지의 땅파는 앞발, 물갈퀴가 달린 양서류의 앞발이 있습니다. 이 모든 특징들은 서로 동일한 구조를 가졌지만 서로 다른 기능을 합니다. 표준 계통발생수는 왜 이렇게 동일한 구조를 가지게 되는가를 보여줍니다. 즉 이들은 이러한 구조를 가진 공통조상을 가진 것입니다. 이것은 계통발생수에 의해서 지지되는 결론입니다. 비록 이 상동기관적인 특징이 이들 종들에서 서로 같은 그룹으로 묶는데 쓰이지는 않지만, 객관적으로 본다면, 이것은 진정으로 놀라운 결과입니다. 왜냐하면, 오직 공통적인 특징(즉, 같은 기능과 구조를 가진)만으로 계통발생수에서 한 그룹으로 묶이기 때문입니다. (cladistic methodology 에 자세한 설명이 있음)
게다가, 화석으로부터 얻은 상호 독립적인 증거들은 많은 이러한 구조들이 다른 구조에서 유래했다는 것을 확인했습니다. 회석기록은, 수각류 공룡과 현대의 조류사이의 중간형태들에 대해서 하나의 일반적이고 연대기적인 진행과정을 보이며 이 과정에서 수각류의 구조가 현대 조류의 구조로 변화됩니다(Carroll 1988; Carroll 1997; Sereno 1999).
이 일련의 과정의 좋은 예로는 에오랩터 (~230 Mya), 헤레라사우리다(the Herrerasauridae, ~230-210 Mya), 케리토사우리다(Ceratosauria, ~220-65 Mya), 알라사우로이디아(Allosauroidea, 180-90 Mya), 데이노초사우리아 (Deinonychosauria, 150-65 Mya), 시조새(~150 Mya), 콘푸시우소니티대(Confuciusornithidae, 145 Mya), 에난티오르니테스(Enantiornithes, 145 Mya-65 Mya), 및 에우오르니텟(Euornithes, 65 Mya-recent) (Sereno 1999)가 있습니다. 그림 3.1.1 은 새의계보에 있어서 4가지 대표적인 중간형태 종의 앞발을 보여주고 있습니다 (Carroll 1988, p. 340; Carroll 1997, p. 309).
반증가능성:
화석기록은 새의 날개가 점차적으로 파충류의 발로 변형되는 연대기적 진행을 보여줄 수도 있습니다. 그러나, 그렇지 않고 그 반대입니다. 게다가 만약 한 유기체의 예상 조상의 원시적인 구조가 합리적으로 현대 유기체의 파생 구조로 변화될 수 없다는 것을 긍정적으로(positively) 증명된다면 아주 강력한 반증이 될 것입니다. 분명하고 가상적인 예를 들어보면, 매우 진지하게, 페가수스와 같은 동물을 발견하는 것이 대진화론적으로 불가능하다는 것입니다. 페가수스는 말과 가장 비슷한 포유류일 것이며 그의 날개는 아마도 파생적인 특징으로 간주될 것입니다. 그러나, 페가수스의 날개는 페가수스 조상의 구조의 변화가 아닙니다. 왜냐하면 페가수스와 말의 조상은 변화가 가능한 구조가 없기 때문입니다(Futuyma 1998, p. 110).
이와 마찬가지로, 우리는 날개와 팔을 동시에 가진 새를 발견하지 못할 것이며, 엽록체를 보유한 연체동물을 발견하지 못할 것입니다, 비록 이러한 구조가 이들 유기체에 매우 유용할 지라도. 이와 마찬가지로 계통수가 구조적인 연속성이 없지만 기능적인 연속성만 있거나 혹은 어떠한 확인된 연속성이 없다면 강력한 진화론의 반증이 될 것입니다. 다른 반증가능성은 예측 3.4을 보십시오
예측 3.2: 분자수준의 상동기관(Molecular paralogy)
상동기관이라는 개념은 유기체의 거시적 구조와 분자수준의 구조에 동일하게 적용됩니다.
확인:
분자수준에서의 상동기관은 매우 인상적입니다. 서로 전혀 다른 기능을 가지는 단백질이 서로 아미노산 서열과 3차원 구조가 비슷한 경우가 많습니다. 자주 인용되는 사례는 라이소자임(lysozyme) 과 α-락탈부민(α-lactalbumin)입니다. 거의 모든 동물들은 라이소자임을 가지고 있습니다. 이것은 방어의 목적으로 분비되어 박테리아 세포벽을 녹이는데 사용되는 단백질입니다(Voet and Voet 1995, p. 381). α-락탈부민은 포유류의 유선에서 유당 합성에 관여하며, 라이소자임과 기능을 매우 다르지만 구조가 매우 유사합니다(Acharya et al. 1989; Voet and Voet 1995, p. 608). 종종 분자 계통발생수로 볼 때 좀더 근본적인 기능을 가진 즉 라이소자임이 좀더 오래된 단백질로 추론됩니다(Prager and Wilson 1988; Qasba and Kumar 1997).
좀더 큰 스케일로 본다면, 이러한 진화론적인 예측의 놀라운 확증은 사카로마이세스 세레비제(Saccharomyces cerevisiae, 이스트, 제빵용 효모)와 예쁜꼬마선충(Caenorhabditis elegans)에서 나왔습니다. 이 두 유기체의 게놈은 최근에 염기 서열이 분석되었습니다(Barrell 1996; Caenorhabditis elegans Sequencing Consortium 1998). 단세포 유기체인 효모에서 사용되는 유전자는 대부분이 모든 생명체가 수행해야 하는 핵심 생화학 기능과 직접적으로 관련되어 있습니다. 진화론적 관점으로 보면 우리는 이 유전자들이 매우 오래된 것이라고 예상할 수 있습니다. 그러므로 우리는 이 유전자의 상당부분이 선충에서도 발견되리라고 기대할 수 있습니다. 이와 반대로 다세포성(multicellularity)과 관련된 유전자는 좀 더 최근에 진화되었을 것입니다. 계통발생적 분석은 이것이 사실임을 보여주었습니다. 선충에 추가된 유전자의 상당수는 진화론적인 예측대로 세포의 핵심기능을 하는 유전자로부터 직접적으로 유래된 것으로 보입니다 (Chervitz et al. 1998).
알려진 진핵세포 유전자의 좀더 광범위한 연구에서도 상동(paradogy)이 자연계에서 매우 흔하며, 진정으로 구조적인 개혁이 극히 드믈다는 것이 보여졌습니다(Rubin et al. 2000). 사이언스의 한 특별호에서 50명이 넘는 연구자들이 노랑초파리(Drosophila melanogaster), 예쁜꼬마선충(Caenorhabditis elegans), 효모(Saccharomyces cerevisiae), 와 인간 게놈의 전체 염기서열의 내용을 검토했습니다. 예쁜꼬마선충(매우 중요한 실험 유기체)에서는 18,000여개의 확인할 수 있는 유전자가 있었으며, 이것의 약 절반은 같은 게놈의 다른 유전자의 복제(duplication)이었습니다. 이와 비슷하게 곤충의 유전자의 70%가 다른 유기체와 공유하며 이것은 대부분의 유전자가 진화를 거쳐서 서로 다른 유기체 서로 다른 기능을 위해서 재사용된다는 것을 나타내고 있습니다. 이러한 사항은 거의 동일한 3차원 구조를 하는 것으로 알려졌기 때문에 과소평가되었음이 분명하며 이러한 유사성은 단지 염기서열의 비교만으로는 검지해 낼 수 없습니다.(단백질, 핵산의 구조적 기능적 많은 중복성에 대해서는 예측 4.1과 4.2에서 좀더 자세히 다룹니다.) 놀랍게도 일부 초파리는 형태학적으로 거의 구분하기 어렵지만 (노랑초파리와 검정초파리와 같이) 유전자의 유사성은 겨우 70%에 달할 뿐입니다(Schmid and Tautz 1997). 최종분석에서 진핵세포의 진화동안 진정한 구조적 혹은 게놈의 혁신은 매우 적었으며, 동일한 유기체 혹은 서로 다른 유기체에서 모두 대부분의 유전자가 약간의 변화만으로 복제되었거나 재사용 되었습니다. 게다가 유기체간의 불일치성의 수준(~30%, 분리된 이후 유전적 진화된 정도를 나타냄)은 분명히, 초파리의 다양한 종의 분리를 일으키는 과정과 같은, 점진적인 소진화과정으로 설명될 수 있습니다.
반증가능성:
최근 진화된 기능을 수행하는단백질은 핵심 기능을 수행하는 단백질과 통계학적으로 유의미한 유사성을 보입니다. 만약 그렇지 않으면 진화론적으로 문제가 됩니다. 게다가, 계통수에서 가까운 곳에 아니라 아주 먼 곳에서 유래한 유전자를 우리가 발견한다면 진화이론과 일관성이 없어지게 됩니다. (즉, 핵심기능을 하는 단백질 보다 더 오래된 유전자라면)(Li 1997; Chervitz et al. 1998)
예측 3.3: 해부학수준의 상사 (Anatomical analogy)
진화론적인 기회주의의 당연한 결과는 상사기관입니다. 상사기관은 서로 다른 종들 사이에서 서로 다른 구조를 가지고 있으나 동일하거나 유사한 기능을 하는 사례입니다. 서로 상이한 종들은 서로 상이한 역사와 서로 다른 구조를 가졌습니다. 만약 서로 상이한 두 종 모두 같은 기능을 진화시켰다면 그들은 새로운 기능을 수행하기 위해 서로 다른 구조를 새로 이용했을 것입니다. 상사는 구조적인 연속성의 원리를 확증해야만 하며, 상사는 예상 조상의 구조로부터 설명되어야 합니다.
확인:
기능적인 상사를 보이는 해부학적 사례는 매우 많습니다. 한가지 사례는 척추동물의 눈과 두족류(오징어가 대표적)의 눈입니다. 앞서 말했지만 미국과 사하라 사막의 식물은 건조한 불모의 지역에서 필요한 동일한 기능을 위해서 서로 다른 구조를 이용한다는 것입니다. 어떤 포유류는 (고래, 매너티, 돌고래), 새(펭귄)와 물고기는 모두 수중환경에서 살고 수영하는 능력을 가지고 있습니다. 그리고 이들은 모두 수중에서 사용할 기능을 위해서 서로 상이한 구조를 이용합니다. 비록 현재는 변화되었지만 이러한 기능을 수행하는 구조들 모두는 그들의 예상선조에서 모두 존재합니다.
반증가능성:
우리는 돌고래, 고래, 펭귄, 혹은 어떠한 포유류 친척들에게서 아가미(물고기의 상사기관이 될 수 있는)를 가진 새로운 종을 발견할 것으로 기대하지 않습니다. 왜냐하면 그들의 가까운 조상은 아가미가 유래할 만한 아가미 혹은 아가미와 유사한 구조가 없기 때문입니다. 아가미가 수중 생활을 하는 포유류나 조류 생명체에도 유용함에도 불구하고, 이것이 대진화론적인 예측입니다. 또한 아래의 분자적 상사기관의 반증가능성도 참고하시길 바랍니다.
예측 3.4: 분자수준의 상사(Molecular analogy)
상동기관처럼 상사기관도 거시적 혹은 분자적 수준에서 모두 나타나게 된다.
확인:
서브틸리신(subtilisin), 카르복시 펩티다제 II(carboxy peptidase II), 와 키모트립신(chymotrypsin)은 널리 알려진 분자적인 사례입니다. 이들 3개 단백질은 모두 세린프로테아제(즉 다른 단백질을 분해시킬 수 있다)입니다. 그들은 같은 기능 활성이 있는 부분에서 서로 동일한 촉매 잔기(catalytic residue)를 가지며 동일한 촉매(catalytic) 메카니즘을 가지고 있습니다. 그러나 이들은 유전자 서열이나 유전자의 구조적인 유사성이 없습니다(Voet and Voet 1995, p. 394).
또 다른 분자적인 사례는 DNA 폴리머라제입니다. DNA 폴리머라제는 DNA 가닥을 복제하는 것을 촉매하는 단백질입니. 즉 이것은 DNA 가닥에 여러개의 뉴클레오타이드를 이어줍니다. DNA 폴리머라제는 쥐의 폴리머라제 β를 제외하고는 모두 구조적으로 유사합니다. 쥐의 폴리머라제 β를 제외하고는 모든 DNA 폴리머라제는 발산형 진화와 관련되어 있습니다(Davies et al. 1994; Voet and Voet 1995, p. 1040). 쥐의 폴리머라제 β는 nucleotidyl transferase라는, DNA 사실에 하나의 뉴클레오타이드를 이어주는 반응을 촉매하는 효소와 구조적으로 유사합니다. 쥐의 폴리머라제 β는 분명히 nucleotidyl transferase에서 돌연변이가 일어나서 단지 한 개의 뉴클레오타이드가 아닌 여러개의 뉴클레오타이드를 추가해 이어주는 것으로 진화한 것이 명백합니다(Aravind and Koonin 1999).
반증가능성:
상동이나 상사는 대진화와 진화론적 기회주의 원리에만 의존하는 예측입니다. 생물학적으로 상동이나 상사가 전혀 없는 경우도 있을 수 있습니다. 예를 들어 생명체는 모듈 방식으로 구성되었을 수도 있습니다. 마치, 각 특정 구조는 각각 특정한 기능 한가지 만을 수행하는 대부분의 인류발생적 창조처럼.
예측 3.5: 미최적화된 해부학적 기능
진화론적 기회주의의 또 다른 귀결은 명백히 최적화 되지 않은 기능입니다. 앞서 언급했듯이 유기체는 새로운 기능이 진화함에 기존에 가지고 있던 것을 이용해야 합니다. 그러므로 처음부터 최종 기능이 알려졌다면 아마도 다르게 배치(즉, 좀더 효율적으로)된 구조에 의해서 기능이 수행되었을 것입니다.
"미최적화된 기능"은 구조적인 기능이 열악하다는 것이 아닙니다. 이것은 단지 좀더 효율적인 설계 (과잉 복잡성을 줄이는 설계)를 가진 구조로 최종 기능을 동일하게 수행할 수 있다는 것을 말합니다. 미최적화된 기능을 수행하는 구조들은, 예를들어 가까운 유기체나 화석 기록등으로 계통발생이 결정되는 것과 같이, 만약 증거들에 의해서 이들의 역사가 알려졌다면, 조상 구조의 기회주의적인 새로운 사용에 근거를 둔 점진적이고, 역사적이고 진화론적인 설명이 가능합니다.
확인:
포유류의 위장관은 호흡계를 지나갑니다. 기능적으로는 이것은 미최적화된 것입니다. 우리는 숨을 쉬거나 뭔가를 삼키는 것을 동시에 하는 것이 더 유익합니다. 불행히도, 우리는 그렇게 할 수 없으며, 이것 때문에 우리가 기도가 막혀서 죽을 수도 있습니다. 그러나 이러한 배치에는 훌륭한 진화사적인 이유가 있습니다. 포유류가 진화한 오스테오레피폼(Osteolepiformes, 데본기의 폐어)는 숨을 쉬기 위해서 공기를 삼켰습니다. 나중에 포유류의 조상들은 어류의 후각 구멍을 새로 사용하여 육지에서 숨을 쉬는 기능을 하게 되었습니다. 그리고 때마침 원래 냄새를 맡기 위한 기관이었던 콧구멍이 폐로부터 식도의 반대쪽에 있었던 것입니다(Futuyma 1998, p. 5).
또 다른 해부학적 미최적화된 기능은 포유류의 망막이 앞뒤가 뒤집히게 된 것입니다. 망막의 혈관과 신경이 망막앞에 위치하기 때문에 빛은 우선 광감지 세포에 도달하기 전에 이들을 통과해야 합니다. 맹점은 바로 이러한 신경 세포가 모두 모여서 뇌로 연결되기 위해서 망먁을 통과해야 하기 때문에 생긴 것입니다. 이러한 뒤집힌 망막은 타고난 많은 문제점을 해결하기 위해 척추동물의 눈은 다양하고 복잡한 보조 구조(예를들어, 황점[foveas]과 느리고 좀 더 투명한 무수신경)와 메카니즘을 이용하고 있습니다.
두족류(예를들어, 오징어와 문어)는 포유류의 눈과 동일 메카니즘 원리에 근거한 유사한 형태의 눈을 가지고 있습니다. 그러나 포유류의 눈과는 달리 두족류는 망막의 구조가 매우 다릅니다(즉 그들은 뒤집히지 않았습니다.)(Goldsmith 1990; Williams 1992, pp. 72-74). 그리고 그들은 맹점이 없습니다. 이러한 점은 포유류도 맹점이 없는 눈을 가질 수 있음을 보여줍니다.
미최적화된 기능에 관한 다른 사례들은 Jury-rigged Design FAQ을 보기 바랍니다.
반증가능성:
포유류중에서 위장관과 호흡계가 서로 교차하지 않는 종을 발견하거나 혹은 파충류나 포유류 중에서 맹점이 없는 것을 발견하면 진화론에 대한 긍정적인 반증이 될 것입니다. 이것은 미숙한 설계가, 설사 이 문제를 고치는 것이 그 유기체에 매우 이롭더라도 진화론적인 과정으로 "고쳐"질 수 없기 때문입니다. 진화론적으로 허락된 변경은 기존에 존재하는 것에 대한 상대적으로 작은 변화들 뿐입니다.
만약 미최적화된 구조라고 생각했던 어떤 구조가 실제로는 기능적으로 매우 효율적이라고 발혀지면 이 분류의 항목들중 많은 것들은 무효화 될 수 있습니다. 그러나 대부분의 경우에 특정한 구조, 배치에서 중요한 기능을 발견한다고 해서 기본적인 결론이 바뀌는 것은 아닙니다. 예를들어, 척추동물 망막의 맹점이 실질적으로 매우 중요한 기능을 하는데 필요하거나 아직 알려지지 않은, 육상동물에게 특정한 기능이 있을 수 있습니다. 사실상 일부 빈진화론자들은 맹점을 가진 완벽한, 앞뒤 뒤집힌 척추동물 눈이 육상생활에 필요하고 좀더 충분한 두족류의 뒤집히지 않은 눈이 해저의 희미한 곳에서 더 어울린다고 제안합니다(Bergman 2000). 그러나 그렇다면 '왜 척추동물인 어류는 앞뒤 뒤집힌 눈을 가지고 있는가'라는 새로운 질문이 생깁니다. 물고기에 있어서 이미 더 깔끔하고 좀더 효율적이고 더 복잡한 두족류의 눈이 물속에서 좀 더 잘 기능할 수 있음에도 불구하고 척추동물의 눈은 왜 더 필요없이 복잡한 것입니다. 미최적화 기능 논증은 논박되지 않았으며, 단지 강조점이 하나의 유기체에서 다른 유기체로 옮겨간 것 뿐입니다. 게다가 verted 눈을 가진 경골어류를 발견함으로써 대 진화론 가정을 반증할 가능성은 아직도 존재합니다. 좀더 많은 정보를 원하면 "Suboptimality and Irreducible Complexity"를 보기 바랍니다.
예측 3.6: 분자수준의 미최적화된 기능
미최적화 설계의 원리는 생체분자 구성에서도 적용됩니다.
확인:
최근 인간 게놈 서열의 연구에서 우리는 인간의 게놈의 2%만의 DNA 만이 단백질을 만든다는 것을 발견했습니다(International Human Genome Sequencing Consortium 2001, p. 900). 우리 게놈의 전체 45%는 트랜스포존으로 이루어져 있으며, 이것은 각각 알려진 기능이 없습니다. (상당수가 암이나 유전질환을 일으키는 것을 제외하고는)(Deininger and Batzer 1999; Ostertag and Kazazian 2001). 레트로포존의 한가지인 LINE1은 전체인간 게놈의 17%를 차지합니다 (Ostertag and Kazazian 2001; Smit 1996, IHGSC 2001, p. 879-882). 지금까지 조사된 각각의 특정 Alu 트랜스포존은 기능이 없는 것으로 나타났습니다(Deininger and Batzer 1999). 그러므로 만약 이러한 유전적 구성요소들이 사실상 전체적으로는 충실한 기능을 보일지라도 지나치게 숫자가 많다는 것과 병을 일으킬 수 있다는 점을 감안하면 그들은 생물학적으로 알려진 가장 비효율적인 유전자 일 것입니다.
인간 게놈의 20%는 가유전자입니다. 그들은 각각 전혀 기능이 없습니다. 대표적인 사례로는 GDPH 유전자가 있습니다. 인간에 있어서 오직 기능을 제대로 하는 GDPH 유전자는 단 하나뿐입니다. 그러나 최소한 20개의 GDPH 가유전자가 존재합니다. 생쥐에 있어서는 200개의 가유전자가 있으며 이들중 필요한 것은 하나도 없습니다. 게다가 인간과 쥐에서 모두 20-30개의 cyt c의 가유전자 사이에 1개 내지 2개의 기능이 살아있는 카피(유전자)가 존재합니다(Li 1997, p. 349)..
기능이 있는 단백질을 코딩하고 있는 진핵세포의 유전자의 대부분은 인트론이라고 불리는 코딩되지 않은 부분이 끼워져 있습니다. 인트론은 단백질을 만들기 이전에 제거되어야 합니다. 인트론은 척추동물 유전자의 약 80%를 차지합니다(Voet and Voet 1995, p. 1144). 트랜스포존과 비슷하게 인트론 역시 특별한 존재 목적이 없습니다. (아주 드믄 경우에 기능이 있는 RNA를 위한 유전나 조절이나 코딩에 관여합니다.)
진핵세포의 DNA에 있는 나머지는 예를들어 AAAAAA, CACACA, 혹은 CGGCGGCGG 같은 짧은 유전자 서열이 반복됩니다. 이들 각각은 어떠한 기능도 없는 것으로 보이며, 그렇기 때문에 쓰레기 DNA라고 불립니다. 후생생물(metazoan)은 게놈에서 쓸데 없는 DNA를 제거하는 효과적인 메카니즘이 없는 것으로 보입니다. 한번 DNA가 게놈에 끼어들면 그것을 거기에 남게 됩니다.
원생생물(단세포 생물)마져도 진화론적으로 엉터리 설계의 대상입니다. 두 종의 섬모류 Paramecium aurelia 와 Paramecium caudatum은 실제로 형태학이나 표현형으로는 거의 구분할 수 없습니다. 그러나 앞의 것은 게놈에 유전자가 200,000kb 이하지만, 뒤의 것은 DNA가 9,000,000kb 가까이 된며 이것은 실제 필요한 것의 최소 45배가 됩니다(Li 1997, p. 383). Paramecium caudatum은 단세포 생물이면서도 인간이 가진 게놈의 3배가 많은 것입니다.
쓸데없는 DNA가 많을수록 낭비되는 에너지도 늘어납니다. 하지만 이러한 분자생물학적 사례들은 진화론적인 역사를 확신시키는 것입니다. 좀더 많은 사례를 알고 싶다면 예측 4.3-4.5의 사례들을 살펴보십시오(Li 1997).
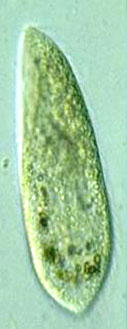
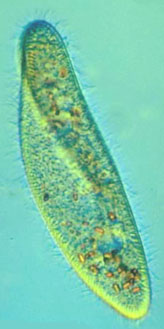 Figure 3.6.1. Paramecium aureliaand Paramecium caudatum
Figure 3.6.1. Paramecium aureliaand Paramecium caudatum 지나간 역사
지나간 역사
 분자 서열 증거
분자 서열 증거